Karbonhidrat Nedir? Glikoz Gibi Şekerler Canlılar İçin Neden Önemlidir?
/old/content_media/4805cc736feafdf96f8a333c78dba2f3.jpg) shutterstock
shutterstock
- Özgün
- Biyokimya
- Beslenme Bilimi
/old/profile_images/0bd861d5b4f09b0f11137736b6902502.jpeg)
/profile/37f26ac92b52d4b1fc6c287a2c379540.jpeg)
/old/profile_images/1bf5e2eaf66a46a83541a8d70be2db9a.jpg)
Bu Makalede Neler Öğreneceksiniz?
- Karbonhidratlar, canlıların temel enerji kaynağı olan ve monosakkaritlerin birleşmesiyle oluşan önemli makromoleküllerdir; glikoz, fruktoz ve galaktoz en yaygın monosakkaritlerdir.
- Monosakkaritler, karbon atomu sayısına ve karbonil grubunun türüne göre sınıflandırılır ve asimetrik karbon atomları sayesinde optik izomerlik göstererek canlıların biyokimyasal çeşitliliğine katkı sağlar.
- Karbonhidratlar, monosakkarit, disakkarit ve polisakkarit olarak çeşitlenir; halkasal yapıları Haworth formülleriyle gösterilir ve bu yapılar canlılarda enerji depolama, yapısal destek ve sinyal iletiminde görev alır.
Vücudumuzu oluşturan yapıların temelinde makromoleküller yer alır. Makromoleküller, monomerik alt birimlerin birbirlerine bağlanmasıyla oluşan, molekül ağırlığı oldukça yüksek olan polimerlerden meydana gelir. Canlı sistemlerindeki en önemli dört makromolekül; karbonhidrat, protein, yağ ve nükleik asitlerdir. Bunlar, hücrenin vazgeçilmez yapıtaşlarıdır; öyle ki, hücreyi bir binaya benzetirsek, binanın yapı taşları bu 4 makromolekülden ibaret olacaktır. Makromoleküller hücredeki metabolik olaylarda, hücre zarının yapısında, sinyal-uyarı iletiminde, yapısal destek sağlamada, canlı için hayati olaylarda gerekli enerjiyi sağlamakta ve bunun gibi birçok yerde görev alırlar.
Makromoleküllerin yapısında en fazla bulunan ve çeşitliliği sağlayan atom olan karbon atomu birçok atomla ya da kendisiyle (en çok da O, H ve N ile) çeşitli bağlar yaparak, yan yana dizilerek zincirler ve yeni bileşikler oluşturur. Bu bileşiklerden birisi de H ve C'nin oluşturduğu hidrokarbon bileşiğidir. Hidrokarbonlar yapılarına oksijeni de katarak canlı için en önemli organik maddelerden birisi olan şekerleri yani karbonhidratları oluşturur.
Karbonhidratlara "şeker" denmesinin nedeni, diğer isimlerinin "sakkarit" olmasıdır (bu kelime, "şeker" anlamına gelmektedir). Şekerler; özellikle de glikoz (İng: "glucose") ve fruktoz (İng: "fructose" veya daha yaygın ismiyle "meyve şekeri") gibi basit şekerler, canlılar için hayati öneme sahiptir. Bu basit şekerler, aynı zamanda monosakkarit ("basit şeker" veya "tek şeker") olarak da bilinir ve canlıların yapısında bulunabilen birçok kompleks şeker molekülünün (nişasta, glukojen, kitin, selüloz, vb.) temelini oluştururlar.
Karbonhidrat molekülü, adından da anlaşılacağı üzere, hidrate olmuş karbon atomları anlamına gelir. "Hidrate olmuş karbon" nedir? Kısaca, su molekülüyle karbonların bileşik oluşturmasıdır; bu bileşiğin formülü de şöyledir: Cn(H2O)n. Oksijen ve hidrojen suda bulundukları 1:2 oranıyla aynı olacak şekilde basit karbonhidrat yapısında da her karbon atomu için bir O ve iki H olacak şekilde bulunur.
/old/content_media/9bde1370814370536a53988dc6186256.jpg)
Bunların hepsi canlılık dahilinde yapıtaşı olarak ve daha önemlisi, enerji kaynağı olarak kullanılmaktadırlar (bir kısmı yapısal olarak da kullanılmaktadır; örneğin kitin isimli karmaşık şeker yapısı, eklembacaklıların dış iskeletlerinde kullanılan ana malzemedir.
Bu şeker molekülleri, farklı sıra ve biçimlerde bir araya gelerek, daha büyük bileşikleri oluşturabilirler. Örneğin yukarıdaki glukoz ve fruktoz bir araya gelerek, sukroz isimli bir diğer şekeri oluşturabilirler:
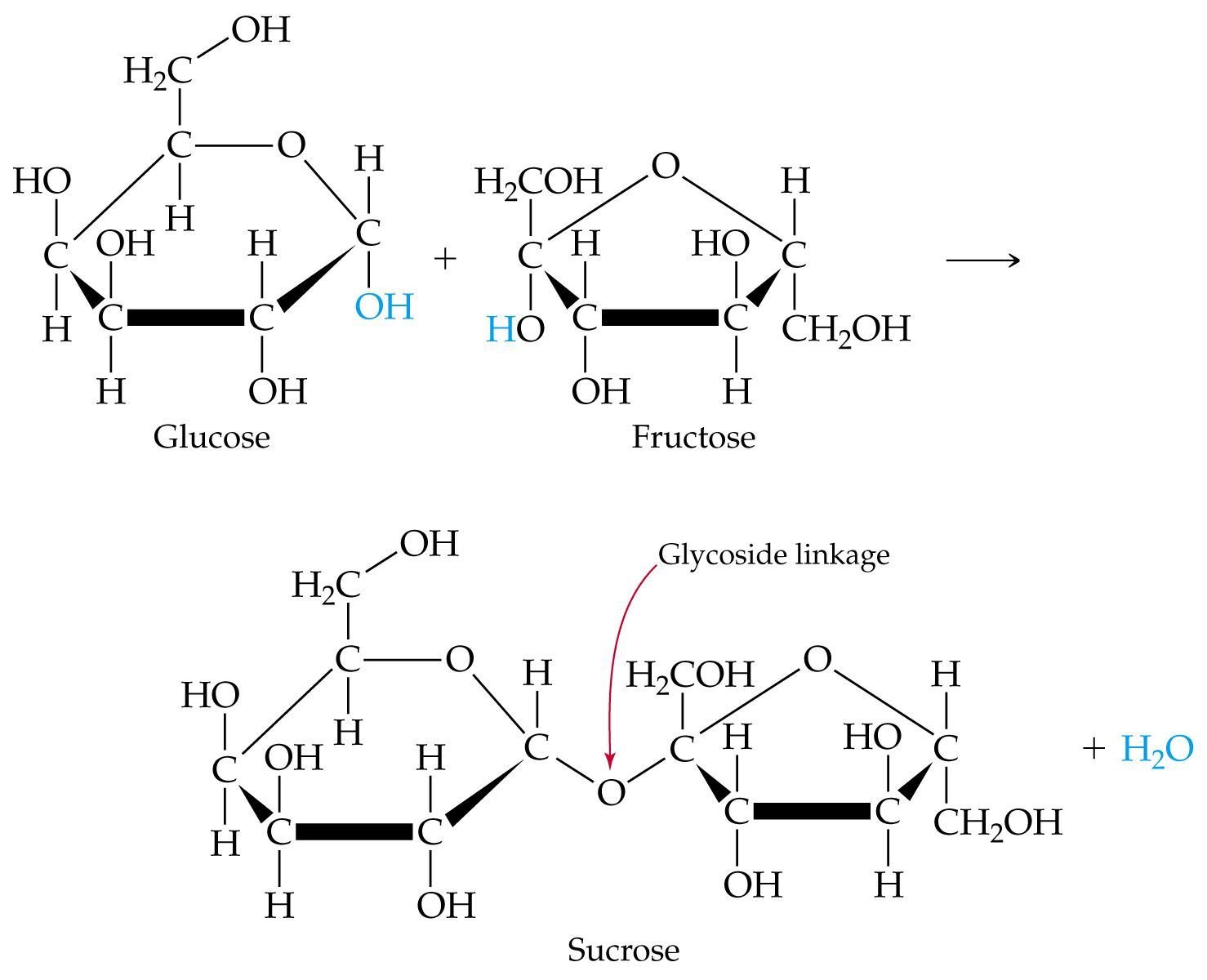
İşte bu şeker molekülleri, daha farklı şekillerde birbirlerine bağlanarak, devasa şeker bileşiklerini oluşturabilirler. Örneğin aşağıda sadece glikozların bir araya gelmesinden oluşan bir glikojen bileşiğini görüyorsunuz:
/old/content_media/0397527d2277fcf3039e8d39c36bc29b.png)
/old/content_media/d7941ca4c22cc26dced9b645553032e3.png)
Doğada en çok bulunan ve canlı sistemlerinde ana yakıt görevi gören karbonhidratlara, hücrenin metabolik faaliyetlerini yerine getirmek için ihtiyaç duyulur: Karbonhidratlar, hücrelere enerjiyi sağlayan makromoleküllerdir. Aslında karbonhidratlar aralarındaki glikozidik bağlar sayesinde diğer makromolekülleri oluşturan bağlardan daha kolay ayrılabildiklerinden bu moleküller esas enerji kaynağı olarak kullanılır; yani yapımları ve yıkımları nispeten diğer pek çok makromoleküle nazaran çok daha kolaydır. Bu sayede yakım ve yıkım oranlarına daha az miktarda enerji harcanmasına bağlı olarak net enerji geliri verimli bir düzeye ulaşmaktadır.
Enerji üretemeyen bir canlının, hayatta kalmak ve üremek için bir aktivite sürdürmesi ve organizasyonunu aktif olarak koruması beklenemez. Dolayısıyla enerji, daha önce de detaylarını gördüğümüz gibi, canlılık için hayati bir kavramdır. Canlılar için bu kadar önemli olan bu molekülleri incelediğimizde, belki de umduğumuzun aksine, pek de sıradışı bir yapı görmüyoruz: 6 adet Karbon atomu (C), 12 adet Hidrojen (H) atomu, 6 adet de Oksijen (O) atomu bir araya gelerek canlıları "canlı" kılan moleküllerden biri olan şekerleri üretebiliyor.
Karbonhidratların Sınıflandırılması
Karbonhidratlar, şeker birimlerinin sayısına ve kimyasal olarak birbirlerine nasıl bağlandıklarına göre çok sayıda gruba ayrılır. Şekerler, nişastalar ve lif bu kategoriler arasındadır. Karbonhidratlar, canlılarda bulunan organik moleküllerin nükleik asitler ve proteinlerden sonra üçüncü büyük grubunu oluşturmaktadır. Organizmada karbonhidratlar monosakkaritler (monomerler) halinde bulunabileceği gibi iki monomer bir araya gelerek disakkaritleri, daha çok monomer bir araya gelerek polisakkaritleri (polimerler) ya da pek çok organik veya inorganik bileşiklerle bağlanarak daha da kompleks yapıları oluştururlar.
Örneğin monosakkaritler, laktoz ve sakkaroz gibi disakkaritleri veya nişasta, glikojeni gibi polisakkaritleri oluşturmak için birbirine bağlanır. Monosakkaritler pürin (adenin,guanin), primidin (timin, sitozin ve urasil) ve fosfatlara bağlanarak nükleik asitleri (DNA, RNA), proteinlere bağlanarak glikoproteinleri, lipitlere bağlanarak glikolipitleri ve diğer bileşiklere bağlanarak glikozitleri, şeker asitleri ve fosfat esterleri gibi karbonhidrat türevlerini oluştururlar.
Evrim Ağacı'nın çalışmalarına Kreosus, Patreon veya YouTube üzerinden maddi destekte bulunarak hem Türkiye'de bilim anlatıcılığının gelişmesine katkı sağlayabilirsiniz, hem de site ve uygulamamızı reklamsız olarak deneyimleyebilirsiniz. Reklamsız deneyim, sitemizin/uygulamamızın çeşitli kısımlarda gösterilen Google reklamlarını ve destek çağrılarını görmediğiniz, %100 reklamsız ve çok daha temiz bir site deneyimi sunmaktadır.
KreosusKreosus'ta her 50₺'lik destek, 1 aylık reklamsız deneyime karşılık geliyor. Bu sayede, tek seferlik destekçilerimiz de, aylık destekçilerimiz de toplam destekleriyle doğru orantılı bir süre boyunca reklamsız deneyim elde edebiliyorlar.
Kreosus destekçilerimizin reklamsız deneyimi, destek olmaya başladıkları anda devreye girmektedir ve ek bir işleme gerek yoktur.
PatreonPatreon destekçilerimiz, destek miktarından bağımsız olarak, Evrim Ağacı'na destek oldukları süre boyunca reklamsız deneyime erişmeyi sürdürebiliyorlar.
Patreon destekçilerimizin Patreon ile ilişkili e-posta hesapları, Evrim Ağacı'ndaki üyelik e-postaları ile birebir aynı olmalıdır. Patreon destekçilerimizin reklamsız deneyiminin devreye girmesi 24 saat alabilmektedir.
YouTubeYouTube destekçilerimizin hepsi otomatik olarak reklamsız deneyime şimdilik erişemiyorlar ve şu anda, YouTube üzerinden her destek seviyesine reklamsız deneyim ayrıcalığını sunamamaktayız. YouTube Destek Sistemi üzerinde sunulan farklı seviyelerin açıklamalarını okuyarak, hangi ayrıcalıklara erişebileceğinizi öğrenebilirsiniz.
Eğer seçtiğiniz seviye reklamsız deneyim ayrıcalığı sunuyorsa, destek olduktan sonra YouTube tarafından gösterilecek olan bağlantıdaki formu doldurarak reklamsız deneyime erişebilirsiniz. YouTube destekçilerimizin reklamsız deneyiminin devreye girmesi, formu doldurduktan sonra 24-72 saat alabilmektedir.
Diğer PlatformlarBu 3 platform haricinde destek olan destekçilerimize ne yazık ki reklamsız deneyim ayrıcalığını sunamamaktayız. Destekleriniz sayesinde sistemlerimizi geliştirmeyi sürdürüyoruz ve umuyoruz bu ayrıcalıkları zamanla genişletebileceğiz.
Giriş yapmayı unutmayın!Reklamsız deneyim için, maddi desteğiniz ile ilişkilendirilmiş olan Evrim Ağacı hesabınıza üye girişi yapmanız gerekmektedir. Giriş yapmadığınız takdirde reklamları görmeye devam edeceksinizdir.
Karbonhidratlar, potansiyel olarak aktif aldehit ya da keton grubu taşıyan polihidroksi alkoller olarak tanımlanmaları yanında, polihidroksi aldehit ya da ketonlar olarak ta tanımlanmaktadır. Karbonhidratlar, CnH2nOn kapalı formülü ile gösterilirler. Kapalı formül parantez içine alındığında, formül birbirinden bağımsız gibi görünen iki parçaya bölünür. Karbon ve su elementlerinden oluştuğu görülür (Latince "Hidrat"). Bu maddeler, formülde görüldüğü gibi karbon ve sudan oluştukları için, "karbonhidrat" olarak adlandırılırlar.
Karbonhidratlar genel olarak her ne kadar CnH2nOn kapalı formülü ile gösterilse de, her karbonhidrat bu formüle uyum göstermez. Örneğin, bir disakkarit olan sükroz (glikoz+fruktoz) C12H22O11 kapalı formülü ile gösterilmekte olup, formülde görüldüğü gibi karbon, hidrojen ve oksijen oranları n, 2n, n şeklinde değildir. Diğer yandan, bu formüle uyum gösteren her bileşik karbonhidrat da olmak zorunda değildir. Örneğin laktik asit C3H6O3 kapalı formülü ile gösterilmesine rağmen karbonhidrat yerine bir organik asittir. Tüm bunlara karşın, genel bir kural olarak karbonhidratların CnH2nOn kapalı formülü ile temsil edildiği ve hidrojenin (H) oksijene (O) oranı genellikle 2/1 olduğu söylenebilir. Bu formülün en iyi temsilcisi, glikozun kapalı formülü C6H12O6'dır ve hidrojenin oksijene oranı 2/1'dir.
Karbonhidratlar farklı şekillerde sınıflandırılmaktadır. Özet bir sınıflandırma şöyle yapılabilir:
- a) Karbon sayısına göre:
- Triozlar (Örn. aldotrioz ve ketotrioz)
- Tetrozlar (Örn. treoz ve eritroz)
- Pentozlar (Örn. riboz, deoksiriboz, arabinoz, ksiloz, liksoz, ribuloz, ksiluloz)
- Heksozlar (Örn. glikoz, alloz, altroz, mannoz, guloz, idoz, galaktoz, taloz vd.)
- Heptozlar (Örn. sedoheptuloz, mannoheptulose vd.)
- Oktozlar (Örn. 3-manno-oktuloz vd.)
- b) Monomer sayısına göre:
- Monosakkaritler (Örn. glikoz, riboz, treoz vd.)
- Glikoz: Vücudun temel enerji kaynağıdır.
- Fruktoz: Şekerlerde bulunan, tatlı bir şekerdir.
- Galaktoz: Laktoz ile birlikte, yaygın olarak süt ve süt ürünlerinde bulunan bir şekerdir.
- Disakkaritler ve Oligosakkaritler (Örn. sükroz, laktoz, maltoz vd.)
- Sukroz: Çay şekeri veya kristal şeker olarak da bilinen şekerdir; glikoz ve fruktozdan oluşur. Şeker kamışından üretilen şeker sukrozdur.
- Laktoz: Süt şekeridir ve inek sütünün %4-5 kadarını oluşturur. Glikoz ile galaktozdan oluşur.
- Maltoz: Malt şekeri olarak bilinir ve nişastanın amilaz enzimiyle sindirilmesi sonucu oluşur. İki glikozun bir araya gelmesiyle oluşur.
- Polisakkaritler (glikojen, nişasta, kitin, selüloz vd.)
- Nişasta: Bitkilerde bulunan uzun glikoz zincirleridir. Sindirmesi kısmen daha kolaydır.
- Glikojen: Hayvanların özellikle de kas ve karaciğer dokularında depolanan glikoz zincirleridir. Genellikle kesim sonrasında çok az depo glikojen kalmasından ötürü besin değeri olan bir şeker türü olarak görülmez. Bir hayvan türü olan insan vücudu da fazladan aldığı glikozu kas ve karaciğerde glikojen olarak depolar.
- Selüloz: Yeşil bitkilerin hücre duvarlarında bulunan şeker zinciridir. Beslenme biliminde "fiber" olarak söz edilen zincir budur. İnsanlar selülozu (fiberi) sindiremez; çünkü evrimsel süreçte 20 yaş dişleri ve apandiks organları körelmiştir. Ancak yine de selüloz tüketmenin, sindirim sistemi için birçok faydası vardır: Örneğin daha hızlı tok hissetmenizi sağlar, daha sağlıklı bir sindirim kanalına sahip olmanızı mümkün kılar ve yüksek fiberli diyetlerin doğrudan veya dolaylı olarak kalp hastalıklarını azalttığı ve bazı kanser türlerine yakalanma riskini düşürdüğü düşünülmektedir.
- c) Aldehit ya da keton grubu taşımalarına göre:
- Aldozlar (Örn. treoz, riboz, alloz vd.)
- Ketozlar (Örn. fruktoz, eritruloz vd.)
Kısaca her şey kimyasal bağlar ve bunların tipleri ile ilgilidir. Aynı temel bileşikler (ya da "hayat molekülleri"), farklı kimyasal bağların etkisi altında farklı yapılara katılabilirler. Zaten hücrelere bakacak olursanız, "hayat molekülleri"nin farklı bileşimlerinden başka bir şey olmadıklarını göreceksiniz. Kısaca farklı bağlar, farklı ürünler demektir. Bu kadar farklı olasılığın, farklı şekillerde yapıya katılması ve milyarlarca yıldır süren Evrim etkisiyle günümüzdeki canlılık çeşitliliğine ulaşırız.
Monosakkaritler ve oligosakkaritler her ne kadar şeker ve şeker benzeri olsa da polisakkaritler ise şeker benzeri olmayan karbonhidratlardır. Bu anlamda monosakkaritler ve oligosakkaritler, "tatlı" maddelerdir. Polisakkaritler hidrolitik olarak parçalandıklarında daha küçük moleküllü polisakkaritlere, yani oligosakkaritlere dönüşür. Parçalanma devam ederse polisakkaritler kendilerini oluşturan monosakkaritlere değin ayrışır.
Şimdi, bahsini geçirdiğimiz her bir karbonhidrat grubunu tek tek inceleyelim.
Basit Şekerler: Monosakkaritler ve Türevleri
Monosakkaritler
Karbonhidratları daha iyi anlamamız için, yapıtaşı olan monosakkaritlerin ne olduğunu bilmemiz gerekir. Monosakkaritler; en basit karbonhidratlardır ve daha basit bileşiklere hidroliz edilemezler. Bu şekerler diğer monosakkaritler ile bir araya gelerek disakkarit, oligosakkarit, polisakkarit gibi polimer yapılardaki şekerleri meydana getirirler. Monosakkaritler, tek bir fonksiyonel karbonil gruptan (aldehit (aldoz) ya da keton (ketozlar)) ve polihidroksi (alkol) gruplarından oluşur. Bu basit bileşikler suda kolay çözünürler fakat polar olmayan çözücülerde (pentan, heksan, benzen) çözünemezler.
Monosakkaritlerin İsimlendirilmesi
Monosakkaritler, içerdikleri karbon atomu sayısına ve yapılarında bulundurdukları fonksiyonel karbonil gruba (C=O) göre isimlendirilirler. Bu bileşenler "şeker" olarak bilinir ve içerdikleri fonksiyonel karbonil grubunun sonuna "oz" eki alarak isimlendirilir. Örneğin insan vücudunda bulunan ve kimyasal açıdan önemli olan monosakkaritler 3-6 karbon içerirler. Bunlar; trioz, tetroz, pentoz, heksozlardır. Bu şekerleri isimlendirirken, içerdiği fonksiyonel karbonil grubu da eklenebilir. 3 karbonlu en basit monosakkaritlerden ve hemen hemen bütün monosakkaritlerin bu bileşiklerden türediği, bir aldotiroz (gliseraldehit) ve bir ketotrioz (dihidroksiaseton) monosakkaritleridir. Aldoheksoz ve ketoheksoz gruplarına, doğada en yaygın D-glukoz ve D-fruktoz monosakkaritleri; aldopentoz grubuna, nükleik asitlerin ve nükleotitlerin yapısına katılan D-riboz (C5H10O5) ve 2-deoksi D-riboz'u (C5H10O4) örnek verebiliriz.
/old/content_media/738bdf2e477839a531f2906f15ea0378.jpg)
Bazı monosakkaritler genelde özel isimleriyle bilinirler; bu isimlendirme, elde edildikleri kaynağa göre verilmiştir. 6C'lu şekerler yani heksozlar; glikoz (üzüm şekeri), fruktoz (meyve şekeri, levüloz) ve galaktoz (süt şekeri) olarak adlandırılan, besin kaynağı bakımından da oldukça öneme sahip monosakkaritlerdir.
Fonksiyonel karbonil grubuna göre isimlendirildiğini dile getirmiştik, bunu açıklayalım: Fonksiyonel karbonil grubu, omurgası dallanmamış karbon atomlarından oluşan monosakkaritlerin karbon zincirindeki bir karbon atomu, oksijen ile çift bağ yaparak karbonil grubunu oluşturur. Karbonil grubu da diğer fonksiyonel gruplar gibi bağlandığı monosakkaritin; molekül şeklini, etkileştiği makromolekülleri, yer aldığı kimyasal tepkimelerin hangileri olduğunu bize açıklar.
Monosakkaritlerin Moleküler Yapıları
Monosakkaritler, çoğu dallanmamış karbon atomlarından oluşan iskelete sahiptirler ve C atomlarıyla birlikte H, O atomları da yapılarında bulunur. Bu karbon zincirindeki C atomları birbirlerine tek bağ ile bağlıdırlar. Yapılarındaki aldehit ya da keton grubunu, C=O grubunun (karbonil grup) pozisyonuna bakarak karar veririz. Aldehitlerde, hidrojene bağlı (-CHO) olan karbonil grup uç kısımda yer alırken, ketonlarda karbonil grup iç kısımdaki bir karbon atomuna bağlı (-C=O) olarak yer alır. Bu grupların dışındaki her bir karbon atomu hidrojene ve hidroksi gruplarına bağlıdır.
C atomu bilindiği üzere en fazla 4 bağ yapar. Bir monosakkarit molekülünün, karbon zincirindeki herhangi bir karbon atomunun yaptığı bu 4 bağın hepsinin farklı atom ya da atom gruplarından oluşmasına asimetrik karbon atomu denir. Asimetrik C atomu içeren bir monosakkarit molekülü, optik özellik gösterir. Optik özellik; glukozun polarize ışığı sağa, fruktozun polarize ışığı sola çevirmesi gibi ışığa yön verme yeteneğidir. Polarize ışığı sağa çevirenler dekstrorotator (+), sola doğru çevirenler ise levorotator (-) adını alır.
/old/content_media/63596c9ea74ff4fece6d9f9bde58af0d.jpg)
Stereoizomerler ve Enantiyomerler
Benzer iki monosakkarit, hem aynı sayıda atom içeriyorsa hem de aynı atom türlerine sahipse ama aslında kimyasal, fiziksel ve fizyolojik olarak birbirlerinden çok farklı durumdaysa, bu moleküllere stereoizomerler denir. Örneğin; monosakkarit moleküllerden galaktoz, mannoz, glikoz ve fruktozun kapalı formülleri aynı olsa da bunlar, kimyasal olarak çok farklıdır ve farklı bileşiklerin yapısına katılırlar. Bir molekül iki çeşit stereoizomerden oluşan, molekülleri birbirlerinin ayna görüntüsü olan ve polarize ışığın yönünü değiştiren bu moleküllere optik izomer denir. Optik izomer terimi, Fischer Projeksiyon Formülü kullanılarak ortaya çıkmıştır. Yapısında asimetrik karbon bulunduran moleküller optikçe aktiflerdir ve polarimetre ile aktifliklerini ölçebiliriz.
/old/content_media/03e713964e94aad629bc27cdadbffca7.jpg)
Monosakkaritlerin asimetrik karbon içerdiklerinden bahsetmiştik. Bir monosakkarit molekülünün içerdiği her asimetrik karbon atomu kadar 2n sayıda optik izomeri - stereoizomeri vardır. n sayısı kadar asimetrik karbona sahip molekülün 2n sayısı kadar da optik izomeri vardır. Örneğin; 4 asimetrik karbon atomuna sahip aldoheksozların 24 = 16 stereo izomerleri, tek asimetrik karbon atomuna sahip gliseraldehit molekülünün de 21=2 stereo izomeri vardır.
/store/product/73faf4c8-8ee3-42e1-ae2f-882974d7d711.jpeg)
Hiç Asimetrik Karbon Atomu İçermeyen Monosakkarit Yok mu?
Tabii ki vardır; en basit şekerlerden biri olan dihidroksiaseton molekülü asimetrik karbon atomu içermez fakat bir diğer en basit yapıdaki gliseraldehit molekülü bir asimetrik karbon atomu içerir ve iki stereoizomeri vardır. Bütün monosakkaritler D-gliseraldehit ve L-gliseraldehitin türevlerinden meydana gelmiştir. Yani monosakkaritler fonksiyonel karbonil grubuna (keton ya da aldehit grubu) en uzaktaki asimetrik karbon atomuna bağlı hidroksi grubunun (OH) konumuna göre D ve L ekini alır; OH asimetrik karbon atomunun solundaysa L-gliseraldehit, sağındaysa D-gliseraldehit adını alır. Bu isimlendirme yine Fischer Formülü'ne göre belirlenmiştir. Bu D ve L izomerleri, enantiyomerleri ifade eder ve gliseraldehit molekülüne benzer tipteki stereoizomerlere enantiyomerler denir.
Enantiyomerlerin stereo izomerlerden farkı şudur: Enantiyomer moleküller birbirinin ayna görüntüsü şeklindedirler ve sadece OH grubunun konumu farklıdır. Diğer bütün özellikleri (hem kimyasal hem fiziksel) birbirleriyle aynı olan moleküllerdir. Örneğin bir çift ayakkabı düşünelim, iki ayakkabı birbirlerinin aynısı olmasına rağmen üst üste geldiklerinde birbiriyle uyuşmazlar, yani sağ ayağımıza ayakkabının sol tekini takamadığımız gibi.
D-glikoz ve L-glikoz birbirlerinin enantiyomerleridir ve enantiyomer moleküller optikçe aktiflerdir. D- glikoz polarize ışığı sağa çevirirken L-glikoz ışığı sola çevirir. D-furktoz ışığı sola çevirirken L-fruktoz sağa çevirir. Bu örneklerle D ve L formlarının birbirine optikçe izomer (birbirlerine zıt yönde ışığı çevirirler) oluşunu anlamış oluruz. Eğer D ve L izomerleri konuldukları solüsyonda optikçe aktiflik göstermiyorlarsa yani solüsyonda D ve L izomerleri eşit miktardaysa rasemik ya da DL karışımı denir. Rasemik karışımda optik etkinlik birbirine eşit ve ters yöndedir
/old/content_media/407517b09bfef5e3d8829b862d675038.png)
Epimerler ve Epimerizasyon
Bir monosakkaritin karbon zincirindeki karbonlar, fonksiyonel karbonil gruptan (aldoz ya da ketoz grubunu oluşturan C atomundan) başlayarak zincirin sonuna doğru numaralandırılır. Kapalı formülleri aynı fakat düz zincir formüllerinde 2, 3 ve 4 numaralı karbonları farklı olan sekiz adet D-aldoheksoz (6 karbonlu aldehit grubu) molekülleri vardır.
Stereoizomer monosakkaritlerde, yalnızca tek bir karbon atomuna bağlı OH molekülünün yönü hariç diğer bütün karbon atomlarına bağlı grupların aynı konumda olmasına epimer denir. D-glukoz ve D-galaktoz monosakkaritlerinde 4 numaralı karbona bağlı olan OH grubunun konumu her ikisinde de farklıdır. D-glukoz ve D-mannoza baktığımızda 2 numaralı karbona bağlı olan OH grubunun konumundaki farklılığı görürüz ve bu bileşikler birbirinin epimeridir. Fakat galaktoz ve mannoz birbirinin epimeri olamaz; çünkü bu iki moleküde birden fazla OH'ın konumunda farklılık vardır. Epimerler birbirine dönüşebilir ve bu epimeraz enzimi ile gerçekleşir. Bu olaya epimerizasyon denir.
/old/content_media/9fa458403fb3b2477863886642ef2db1.png)
Anomerler ve mutarotasyon
Monosakkarit molekülü sulu ortamda çözündüğünde; hem alkol grubu yani hidroksi grubu (OH) hem de C=O (karbonil grubu) içerdiği için bu iki ayrı ucu bir araya gelip reaksiyona girerek halkasal yapı meydana getirir. Glikoz molekülü halkasal yapıya dönüşürken, düz zincirindeki fonksiyonel karbonil (aldehit) grubunun C atomuyla (1 numaralı karbon), 5 numaralı karbonun OH grubuyla (alkol grubu) yani alkolle bağ kurar. Bu 6 karbonlu halkasal yapı tıpkı piran halkasına benzediği için bu yapıya piranoz halkası denir. Piranoz halkası oluşurken, fonksiyonel karbonil (aldehit) grubunun C atomuyla, alkol grubu arasındaki reaksiyon sonucu hemiasetal denen yapılar oluşur. Asetal; bir aldehit grubu ile iki alkolün reaksiyonu sonucunda oluşturduğu üründür. Bir aldehit grubu eğer iki alkol yerine bir alkolle reaksiyona girerse hemiasetal ürünü oluşur.
Monosakkaritler molekül içi hemiasetal oluşurtururlar; fakat bu onlara özgü değildir. Örneğin bir ketoheksoz olan fruktoz monosakkaritindeki keton grubu ile alkollerin reaksiyonu sonucunda hemiketal ürünü oluşur. Fruktoz da glikoz gibi halkalı yapı oluşturacağı zaman keton grubunun C atomu ile (2 numaralı karbon), 5 numaralı karbon atomunun OH grubu (alkol grubu) bağ yapar ve furan halkasına benzeyen 5 karbonlu furanoz halkasını meydana getirir. Örneğin; heksoz grubu olan glikoz ve fruktoz monosakkaritlerini sulu ortama koyduğumuzda %99 halkalı yapı meydana getirirler ve bunlar sırasıyla glikopüranoz, fruktofuranoz (en yaygın furanoz halkalı anomer ß-D-fruktofuranoz) formunda halkaları oluştururlar. Bu oran, şekerin yapısına, pH'ına ve sıcaklığa bağlıdır. Ayrıca pentoz grubundan riboz ve 2-deoksiriboz da furanoz halkası oluşturur. Şunu da belirtmek gerekir sadece 5 ya da daha fazla C atomu içeren aldehit grupları piranoz halkası oluşturur.
/old/content_media/9cc391be6bd34617a66c853f8a0514cc.jpg)
Monosakkaritlerin halkasal yapısının gösterilmesinde Haworth Formülleri kullanılır. Bu halkasal yapı da karbon atomları gözükmez, monosakkaritin 3D görüntüsüne ulaşılır. Gerçek görüntüleri düzlemsel olmayan yani beşgen veya altıgen bir halka görüntüsünde olmayan monosakkaritler kayık, sandalye ve zarf gibi şekillere benzerler. Fischer formülü gerçeğe uymayacak kadar karbon oksijen atomlarını uzattığı için gerçek monosakkarit yapısından uzaktır. Haworth formülü ise halkadaki grupların uzaysal konumları doğru olarak göstermese de atomların birbirine olan uzaklıkları orantılı olduğu için gerçeğe daha yakın bir formüldür.
Monosakkarit molekülünün keton ve aldehit grubundaki C atomu; düz zincir halindeyken asimetrik değilse, sulu ortamda halkasal yapıya dönüştükten sonra (hemiasetal ya da hemiketal oluşturduktan sonra) asimetrik özellik kazanıyorsa bu karbon atomuna anomerik karbon denir. C atomu anomerik karbona dönüştüğünde 2 izomer meydana getirir ve bu izomerler sırasıyla 𝒂 ve ß izomerleridir. Oluşan halkalarda anomerik karbon atomuna bağlı OH grubu halkanın altındaysa 𝒂 izomeri, halkanın üstündeyse ß izomeridir. Hemiketal veya hemiasetal oluşmasından dolayı karbon atomlarının yerinin farklılık göstermesiyle, stereo izomerler meydana gelir ve bunlara anomer denir. Örneğin Haworth formülüne göre, 𝒂 D-glikoz molekülünün, 1 numaralı karbon atomuna bağlı olan OH grubu halkanın altında, ß izomerinin OH grubu ise halkanın üstünde yer alır.
/old/content_media/381c369e718d53a376c086f0f1e541ca.jpg)
Sulu ortamda sadece bir anomer varsa örneğin 𝒂 varken zamanla diğer ß anomeri de ortamda oluşur. Bunun nedeni şudur: 𝒂 ve ß anomerlerinin sulu ortamda dengelenene kadar birbirlerine dönüşmesidir. Örneğin; D-glikoz molekülü sulu ortama konulduğunda polarize ışığı sağ yönde çevirir; fakat gittikçe sağ yönde çevirme açısı azalır, sonunda sabit hale gelir. Bu olayda 𝒂-D-glikoz ve ß-D-glikoz anomerleri dengelenene kadar birbirine dönüşür ve böylece aynı optik özelliklere sahip olan dengeli bir karışım (rasemik karışım) elde edilmiş olur. Bu karışımın: üçte biri 𝒂 -D-glikoz, üçte ikisi ß-D-glikoz ve çok az miktarı glikofuranozdan oluşur. Anlatılan birbirine dönüşüm olayı spontan halde ya da bir mutarotaz enzimi ile olur, bu olaya mutarotasyon denir. Mutarotasyon polarimetre ile gözlemlenebilir. Bu olay sadece glikozda değil diğer monosakkaritlerde de gözlemlenebilir.
/old/content_media/16e7dd4ab8de7a82184a1fe0a98ee24b.png)
Kanımızda ve dokularımızdaki karbonların ana kaynağı, yaşamımız için eşsiz bir değeri olan ve bitki fotosentezinin ana ürünü olan glikoz; yediğimiz meyvelerde bulunup, vücudumuzda kullanabilmek için glikoliz olayında glikolize dönüştürülen fruktoz; laktoz sentezinde kullanılan, süt şekeri olarak da bilinen galaktoz; her organizmada bulunan genetik materyalin yapıtaşı, nükleik asitlerin (RNA ve DNA) yapısına katılan riboz; doğada karşımıza en çok çıkan, biyolojik ve fizyolojik açıdan canlı sistemleri için büyük öneme sahip monosakkaritlerdir ve bunlar canlıların yapısına D izomerleriyle katılır. Bu monosakkaritlerin arasında doğada genel olarak serbest halde bulunanlar glikoz ve fruktoz molekülleridir.
Monosakkaritlerin Önemli Türevleri
Canlı organizmalar vücutlarında birçok monosakkarit bulundururlar fakat bunlar haricinde yapılarında, monosakkkaritlerin kendi içinde molekül yapılarını değiştirerek oluşturdukları monosakkarit türevi moleküller de bulunur. Bu moleküllerden birisine yukarıda anlattığımız bir monosakkaritin halka yapısının oluşumunda rol oynayan glikozid molekülünü örnek verebiliriz. Hatırlayacak olursanız, halka yapısı oluşurken monosakkarit molekülü kendi içinde reaksiyona girerek hemiasetal yapılar meydana getirmişti.
Bu yapı oluşurken karbonil grubunun karbonuyla, geri kalan alkol grubunun OH’ı (hidroksi grup) birbirleriyle glikozid bağı kurarak, piranoz veya furanoz halkasını oluşturur. Bu bağ sadece molekül içinde olmayabilir, iki ya da daha fazla monosakkarit molekülleri arasında da kurulabilir (disakkarit, oligosakkarit, polisakkarit molekülleri). Glikozidler bir monosakkarit ve karbonhidrat olmayan molekül arasında da bağ kurabilir. Bu moleküller örneğin fenol gibi bir alkol bileşiği ya da adenin bazı da olabilir. Bu bağ; monosakkaritin anomerik -OH (hidroksi grubu) ile kurulur.
Hem D-riboz molekülünden bir OH çıkarılarak oluşan, DNA yapısındaki 2-deoksi hem de hücre zarının yapısında önemli yere sahip olan glikolipit ve glikoprotein moleküllerinin yapısında L-fukoz (6-deoksi L- galaktoz) ve L-ramnoz (6-deoksi L- mannoz) olarak adlandırılan deoksi şekerleri ve DNA ve RNA'yı oluşturan şeker fosfatları yani riboz-5 fosfat ve 2-deoksi riboz-5 fosfat molekülleri monosakkarit türevlerine örneklerdir. Şeker fosfatları, monosakkaritlerin fosfat ile esterleşmesi (fosforilasyon) sonucu oluşur ve metabolik olaylar bakımından oldukça önemlidir.
/old/content_media/1f1230668186da63a1060e77aca65d63.jpg)
Bakterilerin hücre duvarının yapıtaşı olan peptidoglikan (N-astilmuramik asit) ve böcekler gibi kabuklu hayvanların kitin tabakasının yapısında bulunan amino şekerler, bir monosakkaritin, OH gruplarından birisinin yerine amin grubu (NH2) bağlanarak oluşan bir monosakkarit türevidir.
Canlılarda monosakkaritlerin oksitlenmiş hali yani şeker asitleri bulunur. Bu yapılar, karbonil grubu ve hidroksil grubunun ya da her ikisinin de oksitlenerek karboksil grubuna (COOH) dönüşmesi ile oluşur. Aldozların aldehit gruplarının, zayıf okside edici bir molekülle karboksil grubuna oksitlenmesi sonucunda aldonik asit oluşur. Örneğin; meyve ve bitkilerin yapısında ve insanlar tarafından tüketilen ürünlerin içerisinde bulunan glukonik asit; D-glukoz’da 1 numaralı karbonun oksitlenmesiyle ve bu grubun karboksil gruba dönüşmesi sonucu meydana gelir. Aldozlarda aldehit grubu değişmeden, 6 numaralı karbondaki alkol grubunun karboksil grubuna oksitlenmesiyle üronik asit oluşur.
D-glukozdan meydana gelen bir üronik asit çeşidi glukoronik asit, detoksifikasyon görevi görür ve bu aside bağlanan her molekül suda kolayca çözünür hale gelir. Aldozlar nitrik asit gibi kuvvetli bir okside edici madde ile, aldehit ve alkol grubunun oksitlenmesiyle aldarik asitler oluşur. Askorbik asit yani C vitamini, insanlar, maymunlar ve kobaylar tarafından sentezlenemedikleri için yeterli miktarda alınmazsa iskorbit hastalığına neden olur. Şekerlerin redüklenmesiyle yani indirgenmesiyle, şeker alkoller meydana gelir. Bu tür şekerlerde karbonil grup, hidroksil gruba (alkollere) indirgenmiştir, bunlar polihidrik alkollerdir. Hücre zarını oluşturan fosfolipidlerin yapısında bulunurlar.
Disakkaritler
Monosakkaritlerin asetal yapı oluştururken, glikozidik bir bağ meydana getirdiğinden bahsetmiştik. İki monosakkaridin 1 numaralı karbon atomunun karboksil grubu ve 4 numaralı karbon atomunun da OH grubu arasında kovalent bir bağ olan glikozidik bağ kurularak ve bunun sonucunda bir molekül su açığa çıkmasıyla disakkaritler oluşur.
Disakkkaritlerin adından, kimyasal yapılarını anlayabiliriz: di=iki sakkarit=şeker, yani iki şekerin birleşmesiyle oluşan moleküllerdir. Onları birleştiren aralarındaki bu glikozidik bağ; 1 ve 4 numaralı karbon atomuna bağlı gruplardan oluştuğu için bu bağa α (1-4) glikozidik bağ ya da β (1-4) glikozidik bağ denilebilir.
Glikozidik bağı asitlere karşı dayanıksız, bazlara karşı dayanıklıdır eğer disakkaritleri monosakkaritlerine ayırmak istersek asitli ortamda kaynatmamız gerekir, böylece glikozidik bağ hidrolize olmuş olur. Doğada yaygın bir şekilde bulunan disakkaritlerden, fizyolojik ve biyolojik olarak büyük öneme sahip olanları maltoz, sakkaroz, laktozdur.
En bilindik depo karbonhidratlarından birisi olan ve besinlerden sık sık aldığımız nişastayı, tükürük bezi ya da pankreasımızda bulunan α-amilaz enzimi ile hidroliz edildiğinde maltozlar meydana gelir. Disakkaritlerin en basit yapısından oluşan bu maltoz, iki D-glukoz molekülünün α (1-4) glikozidik bağıyla oluşur, yani iki D-glukoz molekülünün 1 numaralı α-anomerik karbonun karbonil grubuyla, diğerinin 4 numaralı karbonunun –OH grubu arasında glikozid bağın kurulmasıyla oluşur. Eğer maltoz disakkaritini, monosakkaritlerine ayırmak istersek, ince bağırsakta bulunan maltaz enzimini kullanarak maltozları hidroliz edebiliriz ve böylece iki D-glukoz molekülünü meydana getirmiş oluruz.
/old/content_media/ed2993171c3b629e351ee2d3ba6b674c.png)
Maltoz diğer adıyla malt şekeri sakkoroz şekerine oranla o kadar da tatlı olmayan bir şekerdir. Bu şeker gibi iki D-glikoz monosakkaritinden meydana gelen diğer şeker ise sellobiozdur. Bu şekerin maltozdan farkı, iki D-glukozun birbirine β (1-4) glikozidik bağı ile bağlanarak oluşmuş olmasıdır. Serbest olarak bulunmayan sellobiozlar bitkiler için oldukça önemli olan selüloz polisakkaritini meydana getiren şekerlerdir ve memelilerdeki sindirim enzimiyle değil mikrobiyal enzimler ile ancak parçalanabilirler.
Hepimizin bildiği gibi süt ve süt ürünlerinde sık sık karşılaşılan, sadece meme bezlerinden laktasyon ile sentezlenen laktoz şekeri, D-glikozun D-galaktoz ile β (1-4) glikozidik bağ kurmasıyla oluşur. Laktozlar, laktaz adı verilen ince bağırsaklarımızda mukoza hücreleri tarafından üretilen enzim ile bağlarını hidrolize ederek monosakkaritlerine ayrılırlar. Hatta bağırsaklarında laktaz aktivitesi az ya da olmayan kişilerde laktoz intoleransı rahatsızlığı bulunur ve sindirim sistemlerinde laktozlar sindirilemezler ve emilim olmadan ince bağırsaklarında öylece kalır.
Disakkaritler arasında tat olarak en tatlısı, besinlerimizde oldukça sık bulunan şeker pancarı ve şeker kamışından elde edilen ve diğer adıyla çay şekeri olan sakkaroz (sükroz) şekeri, D-glikoz ve D-fruktozdan meydana gelir ve bu iki monosakkarit anomer karbonlarının glikozidik bağ yapmasından dolayı aralarındaki bağ α β (1-2) glikozidik bağdır.
Oligosakkaritler
Doğal olarak ya da selüloz, nişasta ve glikojen gibi polisakkaritlerin kısmi yıkımı sırasında oluşabilen, örneğin polisakkarit sindirimi sırasında açığa çıkan maltotrioz gibi oligosakkaritler, 3 ya da daha fazla (genellikle en fazla 12) monosakkaritin glikozidik bağlarla bağlanarak polimer bir yapı oluşturması ile oluşurlar. Bitkilerin yapısında fazla bulunan raffinoz önemli bir oligosakkarittir ve hidrolizi ile glikoz, fruktoz ve galaktoz meydana getirir. Oligosakkaritler, hücre zarının yapısında bulunan glikoprotein ve glikolipitlerin N- glikozid veya O-glikozid bağ ile yapılarına katılır.
Polisakkaritler
Makromolekül olan karbonhidrat doğada en çok polisakkarit olarak bulunur. Biyolojik olarak canlı hücrelerinin yapısında önemlidirler çünkü organizmalarda karbonhidratların depo edilmesini sağlamak üzere bulundukları gibi, bağ doku ve hücre duvarı gibi yapısal özelliklerine de katılırlar. Yüksek yapılı bitkilerin depo ürünü olan nişasta, böcek dış iskeletini ve mantarların hücre duvarının yapısını oluşturan kitin, bitkilerin hücre duvarının yapısını oluşturan selüloz, hayvansal hücrelerin depo ürünü olan glikojen, bağ dokusunun yapısına katılan hyaluranik asit gibi önemli molekülleri oluştururlar. Aldığı poli eki ile birçok monosakkaritten oluşan karmaşık karbonhidratlar olduklarını anlarız.
Kendisini oluşturan monosakkaritleri açığa çıkarmak için, asitlerle ve spesifik enzimlerle hidroliz edilmesi gerekir. Karbonhidratların temel bileşeni olan D-glikoz diğer şekerlerdeki gibi polisakkaritlerin yapısında da en çok bulunan monosakkarittir, bunun dışında D-fruktoz, D-mannoz, D ve L-galaktoz, D-ksiloz, D-arabinoz gibi moleküllerde bulunur. Polisakkarit yapısında en çok bulunan D-glikozamin, D-galaktozamin,, D-glikronik asit, N-asetil muramik asit ve N-asetil nöyraminik gibi monosakkarit türevlerini de unutmayalım.
Polisakkaritlerin farklı bir isimlendirilmesi de glikandır. Glikanlar yapısında bulunan monosakkaritlere göre 2 ye ayrılır: Aynı tür monosakkarit-monosakkarit türevi içerenler homopolisakkarit, farklı türlerde monosakkarit-monosakkarit türevi içerenler ise heteropolisakkaritlerdir. En çok bilinen polisakkaritlerin de içinde olduğu homopolisakkaritlere örnek olarak; nişasta, glikojen, dekstran, selüloz, inülin ve kitin’dir. Hetropolisakkaritlere örnek olarak; hyaluronik asit, glikoproteinleri glikolipidler verilebilir.
/old/content_media/846cc85356cae3339a7b05af5d1223ca.png)
Depo Polisakkaritleri
Polisakkaritler proteinler gibi belirli molekül ağırlığına sahip değildirler bunun sebebi, hücreye depo edilirler ve hücrenin karbonhidrat mekanizmasında, karbonhidrata ihtiyaç duyulduğunda bunlar enzimlerle hücrenin ihtiyacı kadar hücreye ilave edilir ya da koparılırlar. Bitki hücrelerinde bulunan nişasta ve hayvan hücrelerinde bulunan glikojen polisakkaritleri bu canlıların sitoplazmasında yer alan depo polisakkaritleridir. Nişastada bulunan α-amiloz, amilopektin ve glikojen α-D- glikopiranozlardan meydana gelmiştir ve yalnızca aralarındaki bağ açısından birbirlerinden farklılık gösterirler.
En çok tüketilen; patates, mısır, pirinç, fasulye, buğday gibi yumru, meyve ve tohumların yapısında daha fazla bulunan ve tüm bitkilerin sentezleyebildikleri bir polisakkarittir. Bitkiler için ilginç bir özellik olan nişasta tanelerinin her bitkiye özgü sıralanışıdır. Nişasta bitkilerde yapısal olarak birbirinden farklı α-amiloz ve amilopektin moleküllerinden oluşur. Nişastanın yapısında amiloz %20-30 kadar bulunur. α-Amiloz gergin olmayan bir bitkide dallanma yapmadan düz zincir halinde ve D-glukoz monosakkaritlerinin α (1-4) bağı ile bağlanmasıyla oluşmuştur.
/old/content_media/b2e4405abf95da7d4688173c23330b53.jpg)
Amilazlar suda çözünüp iyodür çözeltisiyle suya mavi renk verirler. Amilozun yanı sıra amilopektin yapısı en çok α (1-4) bağlarından oluşan düz bir zincir ve yan zincirlerle yani D-glikozun 1 numaralı karbonu ve düz zincirde yer alan D- glikozun 6 numaralı karbonlarının yaptığı dallanmalarla oluşmuş bir yapıdır. Amilopektinler suda çözünemez fakat suya mor renk verir. Nişastanın yapısında genel olarak amilopektin daha çok bulunduğu için, nişasta suda çözünemez ve koyu mavi renk verir. Nişastanın yapısındaki α-amiloz ve amilopektin , tükürük bezi ve pankreasta bulunan salgı bezimizden salgılanan α- amilaz ve β- amilaz enzimi ile hidrolize edilir; fakat bu enzimlerle amilaz parçalanarak dekstrin olarak adlandırılan polisakkariti, maltozu ve glikozu açığa çıkarırken; amilopektinin yan zincirlerden dolayı tamamı hidrolize edilemez. Amilopektinin dallanmış zinciri α(1-6) glikozidaz enzimi ile parçalanır ve amilopektin sonuç olarak tamamen monosakkaritlerine parçalanmış olur, bunun sonucunda glikoz ve maltoz açığa çıkar.
En çok karaciğer hücrelerimizde ve iskelet kası hücrelerimizde sentezlenen ve hayvansal bir depo polisakkarit molekülü olan glikojen, D-glikoz monosakkaritlerinden oluşmuştur ve düz zincirindeki D-glikozlar α-(1-4) glikozidik bağlarla, yan zincirdekiler ise α(1-6) glikozidik bağlarıyla bağlanmışlardır. Buradaki dallanma nişasta yapısında bulunan amilopektinden daha kısa ve fazladır. Glikojen hidrolizine benzer bir hidroliz gerçekleştirir. Düz zincir α- amilaz ve β- amilaz enzimi ile hidroliz edilirken, dallanan büyük zincir ise α(1-6) glikozidaz enzimi ile parçalanır.
/old/content_media/251286d97998fc8724d0ec60e4617822.png)
Diğer depo polisakkaritlerine örnekler verecek olursak; Bakteriler, mayalar, küfler ve yüksek yapılı bitkilerin yapısında mannan bulunur. Yine maya ve bakteri hücrelerinde bulunan dekstranlar türe özgü değişik zincir yapılarından meydana gelen D-glikozlardan oluşan depo polisakkaritidir. Bitkilerin yapısında bulunan fruktan polisakkaritleri de D-fruktoz monosakkaritlerinden oluşmuştur.
Yapısal polisakkaritler
Bilindiği üzere bitkilerin hücre duvarının yapısını oluşturan selüloz polisakkariti, biyosferde fazlaca bulunur ve odun (%50’si), ağaç, çimen, pamuk (%100) gibi bitkilerin yapısında en çok bulunan moleküldür ve selüloz bitkilerin hücre duvarlarının hipertonik, hipotonik gibi su durumları gibi çevre şartları için bitkilere sert yapı kazandırır ve zarar görmesini engeller. Selüloz diğer amilaz, amilopektin ve glikojene yapı olarak benzer fakat farklı olarak D-glikoz monosakkaritleri, β-(1-4) glikozidik bağların bağlanması sonucu oluşmuştur, bu bağlardan dolayı memelilerin sindiriminde görev alan enzimler selülozu parçalayamazlar. Geviş getiren hayvanların ve termitlerin sindirim kanallarındaki mikroorganizmalar sayesinde, mantarların çoğu da ürettikleri spesfik enzimler (sellülaz) sayesinde selülozu parçalayabilir. Eğer kuvvetli asitlerle hidroliz edilirlerse sellobioz ve D-glikoz moleküllerini açığa çıkarırlar.
N-asetil β-D glikozaminlerden meydana gelen kitin, arthropodaların ve mollusca üyelerinin dış iskeletini, mantarların da hücre duvarını oluşturur (bazı mantarlarınki selülozdan oluşur) ve selüloza yapısal olarak benzeseler de kimyasal olarak farklıdırlar. N-asetil-D-glikozaminlerin, β-(1-4) glikozidik bağları ile bağlanmasıyla oluşur ve suda çözünemezler, selülozlar gibi omurgalı hayvanlar tarafından parçalanamazlar.
İnsanlarda ve hayvanlarda bağ dokusunda ve eklem sıvısında hyaluronik asit ana bileşen olarak yer alır ve hatta gözün yapısında bile bulunur. D-glukoronik asit ve N-asetil D-glukozaminin β-(1-3) glikozidik bağ ile bağlanması ile bir disakkarit molekül oluşur, oluşan bu disakkarit molekülleri bir araya gelerek, β-(1-4) glikozidik bağlar ile bağlanarak hyaluronik asitleri meydana getirir. Bu polisakkarit bir heteropolisakkarit yapısındadır. Bakterilerdeki sağlam hücre duvarı yapısının peptidoglikanlar sayesinde olduğunu biliyoruz.
/old/content_media/9b704d7d1944f9fcdbbc78e3c543e483.png)
Peptidoglikanlar, monosakkarit türevleri olan N-asetilglukozamin ve N-asetilmuramik asitin β-(1-4) glikozidik bağlarla bağlanmasıyla oluşan heteropolisakkaritlerdir. Bu yapılar; bakteri hücre duvarında peptid bağları ile çapraz bağlanarak polisakkarit zincirlerini sıkıca hücre zarına bağlar, bu sayede hücre suyun osmotik hareketlerinden dolayı parçalanıp zarar görmesi engellenir. Ökaryot canlılar için oldukça önemli, seçici geçirgen bir yapıda olan hücre zarının yapısına katılıp, hücreye kimlik kazandıran yani bir hücrenin diğer hücreyi tanımasını sağlayan heteropolisakkarit yapılar vardır. Bunlar glikoprotein, proteoglikanlar ve glikolipittir.
Heteropolisakkaritler monosakkarit ya da türevlerinin genellikle proteinlerle ve lipitlerle bağlanmasıyla oluşurlar ve yapılarında sülfat ve fosfat gibi ek gruplar içerebilirler. Bir heteropolisakkarit olan glikozaminoglikanlar, uzun zincirli ve tekrar eden disakkaritlerden oluşmuştur. Glikozaminoglikanlari oluşturan disakkarit molekülleri, N-asetilglukozamin ve ya N-asetilgalaktozamin monosakkarit türevlerinden birini içerir. Disakkarit molekülünü oluşturan ikinci molekül ise çoğunlukla glikuronik asittir. Ökaryot hücreler de hücreler arası boşlukları ekstraselüler matriks sıvısı denilen jel, jöle benzeri yapıdaki sıvılar doldurur.
Proteoglikan, glikozaminoglikanlar ve fibrinöz proteinlerinden meydana gelen bu jel sıvı hücrelerin bir arada tutulmasını sağlayarak hücreye direnç ve esneklik özelliği katar. Canlı sistemleri için fizyolojik ve biyolojik öneme sahip olan glikozaminoglikanlar; hyalüronik asit, kondroitin sülfat, keratan sülfat I ve II, heparin, heparan sülfat ve dermatan sülfattır.
Karbonhidratların en önemli özelliklerinden bilgi taşıma görevi ile proteinlerle birleşerek hücrenin bir başka hücreyle iletişiminde ya da hücrenin ekstrasellüler matriks sıvısıyla arasındaki iletişiminde aracı moleküller olarak görev alırlar. Yani bilgi içeren karbonhidratlar protein ve lipitlerle kovalent bağ yaparak glikokonjugat adı verilen biyolojik olarak aktif görev alan moleküllere dönüşürler. Bunların hücrenin birbirini tanıyıp ayırt etmesini sağlamak, immun yanıt oluşturmak, kan gruplarını oluşturmak gibi birçok görevi vardır. Bu yapılardan birisi olan proteoglikanlar, hücre zarının yüzeyinde ve ekstrasellüler matriks sıvısında bulunan moleküllerdir. Glikozaminoglikanların hücre zarının proteinlerinlerine ya da ortama salınan proteinlerle kovalent bağ yapması sonucu oluşurlar. Proteoglikanların yapısındaki glikozaminler proteinlere oranla daha büyükler (proteoglikanların %95'i karbohidralardan oluşur) ve baskınlardır. Proteoglikan molekülleri bağ dokusu hücrelerine sağlamlık ve esneklik, eğilip-bükülme özelliği veren yapılardır.
Glikoproteinler, polipeptid zincirlerine kovalent bağlarla bağlanan karbonhidratlardan oluşur ve örneğin kollajen, immunoglubin, glikoforin, kan grubu hücrelerinin, sitokinlerin, enzimlerin, hormonların, reseptörlerin ve taşıma proteinlerinin yapısında değişik miktarlarda bulunurlar. Hücre zarına birçok özellik kazandıran hücre zarının yüzey glikoproteinlerinin en önemli görevleri, hücre antijeni oluşturması ve hücreler arası iletişimi sağlamasıdır. Bu özellik hücre zarına seçici geçirgenlik sağlar yani yabancı madde veya organizmaların hücreye giriş çıkışları ayırt edilir. Karbonhidrat içeren kısımları; glikoz, galaktoz, mannoz, N-asetilglukozamin, N-asetilgalaktozamin, sialik asit gibi karbonhidratlardan ve türevlerinden oluşur.
Bu monosakkaritlerin oluşturdukları dallanmış zincirlerden meydana gelen oligosakkaritlerin glikoproteinlerdeki sayıları değişkendir. Hücre de zarın yapısında, kanda, ekstraselüler matriks sıvısında ve bazı organellerin (golgi aygıtı, lizozom, salgı granülleri) yapısında yer alırlar. Oligosakkarit zincirleri fosfolipidlerin hidrofilik baş gruplarıyla kovalent bağ yaparak glikolipitleri meydana getirirler. Glikolipitler hücre zarının yapısında glikoproteinlere oranla çok azdır. Glikoz, galaktoz, N-asetil-galaktozamin ve fükozdan meydana gelen oligosakkaritlerden oluşur.
Bir glikoprotein molekülünde miktarca çok fazla oligosakkarit molekülü bulunurken, glikolipitlerde sadece bir oligosakkarit molekülü vardır. Örneğin kan grubu antijenleri, glikolipid yapısında bulunan oligosakkaritlerin, eritrositlerin (alyuvar hücreleri) zarlarındanki lipidlere bağlanmasıyla meydana gelir. Temel kan grubu O antijenidir fakat O antijenine N-asetilgalaktozamin bağlanmasıyla A grubu antijen, N-asetilglikozamin bağlanmasıyla B grubu antijen oluşur. Karbonhidratların hücre zarında fazlaca bulunduğu bölgeye glikokaliks adı verilir. Genellikle ökaryotlarda bulunan bu yapı, bazı prokaryotlarda da görülür. Sülfat ve iyonize karbonil gruplarından oluşan glikokaliks oligosakkaritlerinin negatif yükleri hücre zarını da negatif yapar.
Gıda Kaynakları
Sebzeler, meyveler, tam tahıllar, süt ve süt ürünleri karbonhidratların başlıca besin kaynaklarıdır. Tahıllar, mısır ve patates de dahil olmak üzere bazı sebzeler nişasta açısından zengindir. Meyveler ve koyu yeşil sebzeler çok az nişasta içerir veya hiç içermez, ancak şeker ve diyet lifi sağlar. Mümkün olduğunca tam tahıllı ekmekler, tahıllar, baklagiller, sebzeler ve meyveler dahil olmak üzere diyet lifi yüksek gıdaların seçilmesi tavsiye edilir. Tipik olarak, sebzeler ve meyveler en konsantre lif kaynakları değildir, ancak bunlar önemli mikrobesinlere katkıda bulundukları için teşvik edilmesi gereken önemli gıdalardır. Benzer şekilde, laktoz içeren süt ve süt ürünleri de genellikle lif içermez, ancak bunlar da diyete kalsiyum, D vitamini ve protein kattıkları için önemlidir.
/old/content_media/022b509beb65435782aa7a5cbcd4dd49.jpg) Hürriyet
HürriyetKarbonhidratlar ve Sağlık
Şekerden bağımsız olarak, tüm karbonhidratlar olumsuz sağlık etkileri ile ilişkili değildir. Nişasta gibi bazı karmaşık karbonhidratlar monosakkaritlere parçalanırken, meyveler, sebzeler ve lif bakımından zengin tam tahıllar ve baklagiller gibi diğerleri farklı bir metabolik yol izler. Emilebilen şekerlerin aksine, bazı kompleks karbonhidratlar insan enzimleri tarafından sindirilemez ve kalın bağırsaktaki mikroorganizmalar tarafından parçalanır. Bağırsak mikroorganizmaları, bu kompleks karbonhidratların fermantasyonunun bir yan ürünü olarak kısa zincirli yağ asitleri (SCFA'ları) üretir. Üretilen birincil SCFA'lar, vücut tarafından bir enerji kaynağı olarak kullanılabilen asetat, propiyonat ve bütirattır. Kompleks karbonhidratlar şekerlerden daha doyurucudur, çünkü SCFA'lar enteroendokrin hücrelerden hormonların ve peptitlerin salınımını artırabilir, bu da artan tokluk ile sonuçlanır, böylece yiyecek arama davranışını azaltır.
Aşırı şeker alımının, özellikle fruktoz etkilerinin aksine SCFA'ların, anti-inflamatuar etkileri, insülin sinyalinin baskılanması yoluyla antidiyabetik etkiler, yağ depolanmasının inhibisyonu ve vücut yağı ve ağırlığının azalması dahil olmak üzere birçok sağlık yararına sahip olduğu görülmektedir. Bağışıklık sistemi üzerinden tahriş edici ajanlara karşı vücudun verdiği tepki inflamatuar olarak bilinmektedir ve aşırı şeker alımı inflamatuarı arttırmaktadır. Aşırı şeker alımı, diyabette bir artış ve inflamasyondaki bir artışın aracılık ettiği bilişsel bozulma ile ilişkilendirilmiştir.
Diyabet ,ayrıca düşük dereceli inflamasyon varlığı ile birlikte insülin sinyallemesinin bozulmasını da birlikte içermektedir. Diyabetin patogenezinin bu yönleri, fruktozun vücut üzerindeki etkileriyle paralel görünmektedir. Fruktoz metabolizması karaciğerde yağ asidi oksidasyonunu artırarak ve inflamatuar faktörlerin transkripsiyonunu artırarak inflamasyona neden olabilir. Bu nedenle, karmaşık karbonhidratların sindiriminde bağırsak mikroorganizmaları tarafından üretilen SCFA'lar, yüksek fruktozlu bir diyetin inflamatuar etkilerine karşı bir koruma kaynağı olabilir.
/old/content_media/ede10f217b3ab6f26c93ff884fbc8ab2.jpg) Mehmet Pense
Mehmet PenseSCFA'ların anti-inflamatuar etkileri beyinde de görülmektedir. Bağırsak mikropları, bağışıklık sistemini ve vagus sinirini içeren yollar aracılığıyla merkezi sinir sistemini düzenler SCFA'lar, portal damarın sinir lifleri, bağırsakların enteroendokrin hücreleri, beyindeki glial hücreler ve adipositler dahil olmak üzere birçok yerde bulunan G-protein reseptörlerine bağlanarak hücre sinyalini ortaya çıkarabilir. Bu bağlanma, bir nöroinflamatuar yanıtını baskılar.
Kompleks karbonhidrat metabolizmasının beyin üzerindeki tam etkileri hala belirsizdir. SCFA'ların, şeker ve yiyecek aramanın etkilerini nasıl dengeleyebilecekleri de dahil olmak üzere, beyin ve davranış üzerinde daha doğrudan bir etkiye sahip olmak için kan beyin bariyerini geçip geçmediğini belirlemek için gelecekteki araştırmalar gereklidir.
Şeker, hem kalori girdisi hem de tat nedeniyle ödül sistemlerimizi tetikleyen oldukça lezzetli bir besindir. Fazla alındığında şeker, bu ödül sistemlerini çok güçlü bir şekilde tetikleyerek fazla yemeye neden olabilir. Striatum merkezli besleyici ve tat ödülü ve hipotalamustan gelen homeostatik sinyal, tokluk sinyali vermek için birbirleriyle iletişim kurmada daha az etkili hale gelir. Glikoz ve fruktozu ayrı ayrı incelediğimizde, eklenen fruktoz, glikozdan çok daha fazla sağlık riski ile ilişkilidir. Fruktoz, yiyecek aramayı artırabilir ve yağ üretimine ve depolanmasına neden olabilir. Ayrıca diyabet ve Alzheimer gibi nörodejeneratif inflamatuar hastalıklarla da ilişkili olabilir. Şekere alternatif olarak meyveler, sebzeler, baklagiller ve lif açısından zengin tahıllardaki kompleks karbonhidratlar bağırsak mikropları tarafından fermente edilir ve şekerin enflamatuar etkilerini dengeleyebilen SCFA'lar üretir.
/old/content_media/056ea839a832dc2ba41f2bb15614d5be.jpg) Beyond the Dish
Beyond the DishŞeker, meyvelerde ve süt ürünlerinde doğal olarak bulunur. Yemeklere üretim ve hazırlık aşamasında olduğu kadar sofralarda da şeker eklenir. Bu "ilave şekerler" (veya dış şekerler), yiyecek ve içeceklerin lezzetini tatlandırır ve geliştirir. Şekerler ayrıca yiyecekleri korumak ve viskozite, doku, gövde ve esmerleşme kapasitesi gibi fonksiyonel özellikler sağlamak için kullanılır. Kalorileri yüksektir, ancak vitaminler, mineraller ve diğer hayati besinler açısından düşüktür. Besin Değerleri etiketi, porsiyon başına toplam şeker hakkında bilgi sağlar, ancak şu anda gıdalarda doğal olarak bulunan şekerler ile ilave şekerler arasında ayrım yapmamaktadır.
Karbonhidratlar Nasıl Evrimleşti?
19. yüzyıldan beridir, şekerlerin doğal süreçlerle oluşabileceğini ve ne tür kimyasalların, hangi şartlarda tepkimesinin şekerleri oluşturacağını net bir şekilde bilmekteyiz. Bu konuda, daha 1861 yılında Alexander Butlerow tarafından yapılan deneyler, şekerlerin oluşumuna ışık tutmuştur. Aldehit ismi verilen kimyasalları yüksek sıcaklıkta ve katalizör kimyasallar etkisi altında bırakan Butlerov, formoz tepkimesi adı verilen bir tepkimeyi gerçekleştirerek basit şekerleri yaratmayı başarmıştır.
Deneyde yapılan, basitçe, formaldehit moleküllerinin yaklaşık 100oC sıcaklıkta kalsiyum hidroksit veya kalsiyum karbonat ile katalize etmektir. Bunun sonucunda ilk etapta glikoaldehitler, sonra D-gliseraldehit ve dihidroksiaseton oluşmaktadır. Bu şekilde devam edilen deneyde D-gliseraldehit ve glikolaldehit molekülleri tepkimeye girerek, riboz gibi 5 karbonlu basit şekerleri üretmiştir. Benzer şekilde, D-gliseraldehit ile dihidroksiaseton moleküllerini karıştırırarak D-glikoz ve D-galaktoz gibi 6 karbonlu basit şekerleri yaratmayı başarmıştır. Tüm bu tepkimelerin bir şemasını aşağıda görmek mümkündür:
/evrimagaci.org/dosyalar/fotograflar/ozgun/62/3585_62_07_evow_ch04.jpg)
Bu şekilde sürdürülen deneyler, canlılığın ilk olarak başladığını sonradan keşfettiğimiz, okyanus tabanlarındaki volkanik bacalar ve etrafındaki kimyasalların keşfiyle daha da isabetli sonuçlar vermeye başladı. Çünkü o ortamların yeniden modellenmesi, buralarda canlılık için gerekecek kimyasalların nasıl sentezlenmiş olabileceğini anlamamızı sağladı. Sadece bu bölgelerde, okyanus tabanlarında bulunan kimyasalların deneylerde analiz edilmesiyle, canlılığı oluşturacak tüm kimyasalların yaratılmasının mümkün olduğu gözlendi.
Örneğin 1989 yılında Alman biyolog Egon T. Degens tarafından yapılan bir araştırmada, formaldehit ile kaolin kimyasallarının 100oC gibi bir sıcaklıkta kalsiyum fosfat ile tepkimesi sonucu basit bir şeker olan D-ribozun tamamen doğal biçimde oluşabileceği gösterildi. Bunun haricinde, aynı deneyde, az sonra değineceğimiz adenin gibi nükleotitleri ve fosfolipitleri de üretebilmiştir.
1960 yılında Mayer ve Jaschke tarafından yapılan bir deneyde, %2'lik formaldehit (0.67 M), kalsiyum karbonat bulunan ortamda geridöndürme (İng: "reflux") tepkimesine maruz bırakılmıştır ve bunun sonucunda ribüloz şekeri üretilebilmiştir. 1967 yılında Reid ve Orgel tarafından deney tekrarlanmış ve kalsiyum karbonatın bu formoz tepkimesinde katalizör etkisi gösterdiği doğrulanmış; ancak kağıt kromatografisi yoluyla tespit edilen ribülozun varlığı doğrulanamamıştır. Ancak ikili, yaptıkları deneyde 0.5 M formaldehitin kalsiyum karbonat veya karbonat-apatit varlığında %40 oranında şeker üretebildiğini göstermişlerdir. Öyle ki, 0.01 M kadar düşük molaritelerde bile şeker üretmek mümkün olmuştur. Aynı deneylerde, yaklaşık 24 saat içinde tepkimeden uzaklaştırılması gerektiğini, aksi takdirde süregelen tepkimenin üretilen ürünleri bozmaya başladığı da gösterilmiştir - ki bu durum, okyanus dibindeki volkanik bacalardaki süreğen akıntılarda bu tepkimelerin oluşup, ürünlerin uzaklaştırılmasının avantajlarına işaret ediyor olabilir.
1967 yılında Gabel ve Ponnamperuma, kaolinit, illit ve alüminyum gibi mineral ve elementlerin varlığında, yukarıdaki tepkimelerdeki ortamın aksine, bazik değil de tamamen nötr ortamlarda bile heksoz, pentoz, tetroz ve trioz şekerlerin oluşabildiğini, hem de 0.01 M kadar düşük molaritelerde bile oluşabildiğini göstermişlerdir. 1972'de Cairns-Smith, 0.13 M formaldehit ile geridöndürme tepkimelerini test etmiş ve kuartz, kalsit ve galena gibi çok sayıda mineralin şeker oluşumunu katalizleyebildiğini göstermiştir.
1993 yılında Schwartz ve Graaf, 0.1 M formaldehitin, ısıtılması halinde Dünya'da yaygın olarak bulunan çok sayıda mineralin etkisi altında katalizlenebildiğini ve otokatalitik (kendi kendinin üretimini hızlandıran) formoz tepkimesinin elde edilebildiğini göstermiştir. Bu tepkimenin kalsiyum iyonlarına veya önceki deneylerde gereksinim olduğu sonucuna varılan güçlü bazik bir ortama ihtiyacı olmadığını da göstermişlerdir. Bu tepkime sonucunda oluşan şekerler seçici değildir; ancak Dünya atmosferindeki formaldehitin fotokimyasal sentezi yoluyla pentaeritritole dönüşümü fazlasıyla seçicidir ve bu süreçte oluşan karbonhidrat oluşumu, yaşamın başlangıcındaki şekerlerin inorganik moleküllerden oluşumunu açıklayabilir.
2020 yılında Haas ve arkadaşları; hidroksitler, karbonbatlar, silikatlar, mikalar, zeolitler, killer, olivinler, fosfatlar ve fosfitler gibi muazzam bir mineral çeşitliliği içerisinde, glikoaldehitten şeherlerin mekanokimyasal bir şekilde oluşabileceğini göstermişlerdir. Üstelik bu tepkimeler oldukça kısa sürelerde yaşanmış ve düzenli bir şekilde monosakkaritleri üretmeyi başarmıştır. Mineral bolluğuna bağlı olarak aldozlar, ketozlar ve hatta spesifik şekerlerin üretimi kontrol edilebilmektedir. Su ortamında değil de atmosferik şartlarda yapılan deneylerde, bu sürecin oksijen varlığından da bağımsız olduğu, yani oksijen olsaydı bile tepkimenin devam edebileceği gösterilmiştir. Benzer şekilde, katyon kompozisyonlarına bağlı olarak tepkimenin seçiciliği geniş bir aralıkta değişebilmektedir ve bu da şekerlerin erken Dünya'da oluşabilmesi için potansiyel kimyasal çeşitliliğini yansıtmaktadır. Araştırmanın yazarları, çalışmalarını şöyle özetliyor:
Genel olarak, mekanokimyasal formoz tepkimesi, erken Dünya koşullarında olan jeokimyasal şartlar altında şekerlerin (monosakkaritlerin) oluşabilmesi için sağlam bir sentez yolağı sunmaktadır. Dahası, katı-faz tepkimesi, şekerlerin su olmayan ortamda da oluşabildiğini göstermekte ve bu sayede Dünya-dışı şeker oluşumuna da kapı aralamaktadır. Bu durum, içinde su bulunmayan meteoritlerde şeker bulunduğunu gösteren yakın geçmişteki araştırmaların bulgularını izah etmektedir.
Evrim Ağacı'nda tek bir hedefimiz var: Bilimsel gerçekleri en doğru, tarafsız ve kolay anlaşılır şekilde Türkiye'ye ulaştırmak. Ancak tahmin edebileceğiniz gibi Türkiye'de bilim anlatmak hiç kolay bir iş değil; hele ki bir yandan ekonomik bir hayatta kalma mücadelesi verirken...
O nedenle sizin desteklerinize ihtiyacımız var. Eğer yazılarımızı okuyanların %1'i bize bütçesinin elverdiği kadar destek olmayı seçseydi, bir daha tek bir reklam göstermeden Evrim Ağacı'nın bütün bilim iletişimi faaliyetlerini sürdürebilirdik. Bir düşünün: sadece %1'i...
O %1'i inşa etmemize yardım eder misiniz? Evrim Ağacı Premium üyesi olarak, ekibimizin size ve Türkiye'ye bilimi daha etkili ve profesyonel bir şekilde ulaştırmamızı mümkün kılmış olacaksınız. Ayrıca size olan minnetimizin bir ifadesi olarak, çok sayıda ayrıcalığa erişim sağlayacaksınız.
Makalelerimizin bilimsel gerçekleri doğru bir şekilde yansıtması için en üst düzey çabayı gösteriyoruz. Gözünüze doğru gelmeyen bir şey varsa, mümkünse güvenilir kaynaklarınızla birlikte bize ulaşın!
Bu makalemizle ilgili merak ettiğin bir şey mi var? Buraya tıklayarak sorabilirsin.
Soru & Cevap Platformuna Git-
 7
7
-
 4
4
-
 3
3
-
 2
2
-
 1
1
-
 1
1
-
 1
1
-
 0
0
-
 0
0
-
 0
0
-
 0
0
-
 0
0
- J. Slavin, et al. (2014). Carbohydrates. Advances in Nutrition, sf: 760-761. doi: 10.3945/an.114.006163. | Arşiv Bağlantısı
- Ankara Üniversitesi Açık Ders Malzemeleri. Biyokimya 1. Hafta Ders Notları. Alındığı Tarih: 20 Ocak 2022. Alındığı Yer: Ankara Üniversitesi Açık Ders Malzemeleri | Arşiv Bağlantısı
- G. Wang. (2018). Impact Of Sugar On The Body, Brain, And Behavior. IMR Press, sf: 2255-2266. doi: 10.2741/4704. | Arşiv Bağlantısı
- T. Ası. (1999). Tablolarla Biyokimya Cilt 1. Yayınevi: Nobel Tıp Kitapevi.
- T. Ası. (2022). Tablolarla Biyokimya Cilt 2. Yayınevi: Nobel Tıp Kitapevi.
- A. L. Lehninger. (2013). Lehninger Biyokimyanın Ilkeleri. ISBN: 9786053551393.
- V. Rodwell, et al. (2018). Harpers Illustrated Biochemistry. ISBN: 9781260288421. Yayınevi: Nobel Kitapevi.
- J. B. Reece, et al. (2021). Biyoloji. ISBN: 9786052828335. Yayınevi: Palme Yayınevi.
- A. W. Schwartz, et al. (1993). The Prebiotic Synthesis Of Carbohydrates: A Reassessment. Journal of Molecular Evolution, sf: 101-106. doi: 10.1007/BF00166245. | Arşiv Bağlantısı
- M. Haas, et al. (2020). Mineral-Mediated Carbohydrate Synthesis By Mechanical Forces In A Primordial Geochemical Setting. Communications Chemistry, sf: 1-6. doi: 10.1038/s42004-020-00387-w. | Arşiv Bağlantısı
Evrim Ağacı'na her ay sadece 1 kahve ısmarlayarak destek olmak ister misiniz?
Şu iki siteden birini kullanarak şimdi destek olabilirsiniz:
kreosus.com/evrimagaci | patreon.com/evrimagaci
Çıktı Bilgisi: Bu sayfa, Evrim Ağacı yazdırma aracı kullanılarak 23/07/2026 20:59:07 tarihinde oluşturulmuştur. Evrim Ağacı'ndaki içeriklerin tamamı, birden fazla editör tarafından, durmaksızın elden geçirilmekte, güncellenmekte ve geliştirilmektedir. Dolayısıyla bu çıktının alındığı tarihten sonra yapılan güncellemeleri görmek ve bu içeriğin en güncel halini okumak için lütfen şu adrese gidiniz: https://evrimagaci.org/s/11342
İçerik Kullanım İzinleri: Evrim Ağacı'ndaki yazılı içerikler orijinallerine hiçbir şekilde dokunulmadığı müddetçe izin alınmaksızın paylaşılabilir, kopyalanabilir, yapıştırılabilir, çoğaltılabilir, basılabilir, dağıtılabilir, yayılabilir, alıntılanabilir. Ancak bu içeriklerin hiçbiri izin alınmaksızın değiştirilemez ve değiştirilmiş halleri Evrim Ağacı'na aitmiş gibi sunulamaz. Benzer şekilde, içeriklerin hiçbiri, söz konusu içeriğin açıkça belirtilmiş yazarlarından ve Evrim Ağacı'ndan başkasına aitmiş gibi sunulamaz. Bu sayfa izin alınmaksızın düzenlenemez, Evrim Ağacı logosu, yazar/editör bilgileri ve içeriğin diğer kısımları izin alınmaksızın değiştirilemez veya kaldırılamaz.