Uzun Vadede Seçilimin ve Genotip Farklarının Evrime Etkisi
:sharpen(0.5,0.5,true)/old%2Fcontent_media%2Fdad018197859a21d6e658e085aeaa1bf.jpg) Video Blocks
Video Blocks
:sharpen(0.5,0.5,true)/profile%2F2aacbbe4-281f-4f73-9317-6cfea6fc8bc0.jpeg)
Bu Makalede Neler Öğreneceksiniz?
- Ortalama uyum başarısı, popülasyondaki her genotipin frekansı ile uyum başarısının çarpımının toplamı olarak hesaplanır ve evrimsel değişimlerin temelini oluşturur.
- Seçilim baskısı altında, genotip frekanslarının değişimi matematiksel formüllerle modellenebilir ve bu değişimler evrimin yönünü ve hızını belirler.
- Farklı seçilim türleri (yönlü, ekleyici yönlü, baskın yönlü, çekinik yönlü) genotiplerin uyum başarısını etkileyerek popülasyonun evrimsel dinamiklerini şekillendirir.
Bir önceki yazımızdan hatırlayabileceğiniz gibi, artık uyum başarısının ne olduğunu, evrimi neden etkilediğini ve bir popülasyondaki ortalama uyum başarısının ne anlama geldiğini biliyoruz. Bu yazımızda da bu konu üzerinden giderek çeşitli popülasyonları, farklı genotip dağılımlarına göre analiz etmeyi öğreneceğiz. Böylece farklı seçilim baskıları altında, farklı genotiplerin ne yönlere doğru evrimleşebileceğini tahmin etmeyi ve hesaplamayı öğrenmiş olacağız. Ancak başlamadan önce, ortalama uyum başarısının nasıl hesaplandığını hatırlayalım ve bunun matematiksel anlamını bir örnek üzerinden görelim. Hatırlayacak olursanız ortalama uyum başarısını şöyle hesaplıyorduk:
ῶ = (FAA * ωAA) + (FAB * ωAB) + (FBB * ωBB)
Hatırlayacağınız gibi burada F harfiyle belirtilenler genotip frekanslarını (örneğin AA genotipinin popülasyon içerisinde bulunma sıklığını), omega (ω) işaretiyle gösterilenlerse, o genotipin çevreye uyum başarısını ifade etmektedir. Dolayısıyla her bir genotipin uyum başarısını, o genotipin bulunma sıklığı ile çarparak sonuçları topladığımızda, popülasyonun ortalama uyum başarısını elde etmiş oluruz. Bu tıpkı şu soruyu çözmeye benzer:
Bir grup çocuğun oluşturduğu bir popülasyonda, çocukların %10'u 1.2 metre, %60'ı 1.5 metre, %30'u 1.8 metre boy uzunluğuna sahiptir. Bu popülasyonun ortalama boy uzunluğu ne kadardır?
Bu hesabı yapabilmek için çocukların toplam sayısına gerek bile yoktur. Tek yapmamız gereken, tıpkı yukarıdaki uyum başarısı formülünde olduğu gibi, boy uzunluklarını (belli bir genotipin uyum başarısına karşılık gelmektedir), o boy uzunluğunun bulunma sıklığı (frekansı) ile çarpıp sonuçları toplamaktır. Bunu yaptığımızda, şöyle bir işlemle karşılaşırız:
ῶ = (0.1 x 1.2) + (0.6 x 1.5) + (0.3 x 1.8)
ῶ = 1.56
Yani böyle bir popülasyonun ortalama boy uzunluğu 1.56 metre olacaktır. Zaten bireylerin çoğunun (%60) boyu 1.5 metre olduğu için, ortalama da bu sayıya epey yakın olmaktadır. Bu şekilde de hesabımızı kısmen test edebiliriz. İşte aynı yöntemle, popülasyonların ortalama uyum başarısını da hesaplamak mümkündür.
Evrimsel biyologlar, bu formülü tam tersine de kullanabilmemiz için ufak tefek oynamalar yapmışlardır. Bunlar basit, lineer işlemlerdir ve bu sayede, verili bir seçilim baskısı altında, bir sonraki popülasyonda doğrudan A alelinin frekansını (p) veya a alelinin frekansını (q) hesaplayabiliriz. Her seferinde yukarıdaki formülle uğraşmamak için, bu formülleri de öğrenmekte fayda görüyoruz.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F61045982_coffee4jpg.jpg)
Seçilim Baskısı Altında Evrim
Şimdi, sadece 1 nesil geçtikten sonra, her bir genotipin frekansındaki değişimi nasıl ifade edeceğimizi öğrenelim:
FAA = (p2 x ωAA) / ῶ
FAB = (2pq x ωAB) / ῶ
Aslında maddi destek istememizin nedeni çok basit: Çünkü Evrim Ağacı, bizim tek mesleğimiz, tek gelir kaynağımız. Birçoklarının aksine bizler, sosyal medyada gördüğünüz makale ve videolarımızı hobi olarak, mesleğimizden arta kalan zamanlarda yapmıyoruz. Dolayısıyla bu işi sürdürebilmek için gelir elde etmemiz gerekiyor.
Bunda elbette ki hiçbir sakınca yok; kimin, ne şartlar altında yayın yapmayı seçtiği büyük oranda bir tercih meselesi. Ne var ki biz, eğer ana mesleklerimizi icra edecek olursak (yani kendi mesleğimiz doğrultusunda bir iş sahibi olursak) Evrim Ağacı'na zaman ayıramayacağımızı, ayakta tutamayacağımızı biliyoruz. Çünkü az sonra detaylarını vereceğimiz üzere, Evrim Ağacı sosyal medyada denk geldiğiniz makale ve videolardan çok daha büyük, kapsamlı ve aşırı zaman alan bir bilim platformu projesi. Bu nedenle bizler, meslek olarak Evrim Ağacı'nı seçtik.
Eğer hem Evrim Ağacı'ndan hayatımızı idame ettirecek, mesleklerimizi bırakmayı en azından kısmen meşrulaştıracak ve mantıklı kılacak kadar bir gelir kaynağı elde edemezsek, mecburen Evrim Ağacı'nı bırakıp, kendi mesleklerimize döneceğiz. Ama bunu istemiyoruz ve bu nedenle didiniyoruz.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fpublic%2Fimages%2Fmisc%2Ffeed-support-3.png)
FBB = (q2 * ωBB) / ῶ
Bu 3 formül de, en yukarıda verdiğimiz formüle uyan bir popülasyonun, 1 nesil geçtikten sonra genotip frekanslarının dağılımını vermektedir. Dikkat ederseniz, genotip frekansları p2, 2pq ve q2, o genotiplere ait uyum başarısıyla çarpılmakta ve sonuç, ortalama uyum başarısına bölünmektedir. Böylece o tekil frekansın 1 nesildeki seçilim baskısı altındaki değişimi hesaplanabilir.
Bu 3 denklemde sağ tarafları topladığınızda 1'e eşit olduğunu görürsünüz. Çünkü her birinin pay kısmında olan ifadelerin toplamı, zaten ῶ değerine eşittir. Dolayısıyla toplama yapıldığı zaman pay ile payda birbirine eşit olacaktır. Bu da 1'e eşitlik demektir. Böyle olmak zorundadır, çünkü denklemin sol tarafında genotip frekansları vardır. Unutmayınız ki bir popülasyondaki alellerin, genlerin, genotiplerin, fenotiplerin veya neyin olursa olsun o şeyin frekansları toplamı daima 1'e eşit olmak zorundadır. Dolayısıyla denklemin sol taraflarının toplamı da, sağ tarafların toplamı gibi 1'e eşittir. Böylece yukarıdaki eşitliklerin doğru olduğunu anlarız.
Şimdi, tek tek alelleri birbirinden ayırabiliriz. Bunun için, önceki yazılarımızda yaptığımız gibi, basitçe alelleri sayacağız. Ancak şu anda elimizde sayısal değerler olmadığı için ayrı alellerin seçilim baskısı altında 1 nesil sonra nasıl değişeceklerinin denklemini doğrudan verelim:
p' = [(p2 x ωAA) / ῶ] + [1/2 x (2pq x ωAB) / ῶ]
Karmaşık gibi görünmesi korkutmasın, çok çok basit bir işlem yaptığımız: Bir sonraki nesildeki A frekansını (p' ile gösteriliyor) bulabilmek için, iki adet terimi birbiriyle topluyoruz. Bu terimlerden ilki, doğrudan homozigot baskın olan AA genotipinin frekansı. Çünkü bu genotipin tamamından A aleli gelecek. Toplamdaki ikinci terim ise, heterozigot olan AB genotipi frekansının yarısıdır. Çünkü heterozigotların yarısı A alelini verebilecektir. Bu formülü biraz daha sadeleştirerek şöyle yazabiliriz:
p' = [(p2 x ωAA) + (pq x ωAB)] / ῶ
Burada tek yapılan, yukarıdaki formüldeki toplamada, ikinci terimde yer alan 1/2 ile 2 çarpımını sadeleştirmek ve ortak olan paydaları birleştirmektir.
Çekinik homozigotu da aynı şekilde hesaplayabiliriz, çünkü yine yarısı heterozigottan gelecektir, diğer kısmı ise tamamen homozigot çekinik genotipten gelecektir.
q' = [(q2 x ωBB) + (pq x ωAB)] / ῶ
Benzer bir şekilde, basit bir hesapla her bir nesilde p ve q frekanslarının ne kadar değişeceğini de gösterebiliriz. Çünkü unutmayın ki yukarıda verilen p' ve q' frekansları, 1 nesil geçtikten sonraki frekanslardır. Ancak arada frekansın ne kadar değiştiğini, bu yeni frekans ile önceki frekansı birbirinden çıkararak bulabiliriz. Değişim, matematikte genellikle Δ (delta) ile gösterilir. Bu durumda
Δp = p' - p
Δp = {[(p2 x ωAA) + (pq x ωAB)] / ῶ} - p
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fpublic%2Fimages%2Fdyn%2Fc11ce4d98b62f2e4ff05cf4f7ad51fa8.png)
Buraya kadar sadece p' ifadesini yukarıdaki formülden çekerek yerine yerleştirdik. Fazladan parantezler sadece netlik ve neyin neye bölündüğünün, hangi işlemin önce yapılması gerektiğinin ifadesi için eklenmektedir. Yukarıda en son verdiğimiz çıkarma işlemi yapılacak ve sadeleştirilecek olursa, şöyle bir ifadeyle karşılaşırız:
Δp = (p/ῶ) x [(p x ωAA) + (q x ωAB) - ῶ]
Aynı şekilde, q frekansının değişimi de hesaplanabilir. Ara basamakları atlayarak yazdığımızda, sonuç şöyle olacaktır:
Δq = (q/ῶ) x [(p x ωAB) + (q x ωBB) - ῶ]
İşte bu 2 formül, doğrudan seçilimin evrime etkisini verir. Çünkü evrimin, bir popülasyondaki gen frekansının değişimi anlamına geldiğini hatırlayınız. Yukarıdaki 2 formül de, 2 ayrı alelin her bir nesilde, belli seçilim baskıları altında (ωAA, ωAB, ωBB) ne kadar değişeceğini göstermektedir. Yani yukarıdaki iki formül, seçilim baskısı altındaki evrimin formülüdür!
Bazı Analizler
Bu noktadan sonra bazı analizler yapacak kadar bilgi edinmişiz demektir. Öncelikle, farklı genotiplerin farklı uyum başarıları etkisi altında nasıl değişeceğini öğrenelim.
Evrim, gizemli amaçları olan, düz mantığın ötesinde bir bilinçle çalışan, ekstrem becerileri olan bir süreç değildir. Yalın ve basit bir doğa yasasıdır ve tüm canlılar bu yasalara boyun eğmek zorundadırlar (belki insan kısmî bir istisna sayılabilir). Dolayısıyla bir popülasyonun matematiksel analizi yapılırken, hesabın tekdüze oluşu bireyi caydırmamalıdır. Bunu tek bir soruyla gösterebiliriz:
Eğer ki elinizde homozigot baskın, heterozigot ve homozigot çekinik özellikleri taşıyan, sıradan bir canlı popülasyonu varsa ve bu genotiplerin uyum başarısı sırasıyla w11, w12, w22 ise ve bu üçlü arasında şöyle bir ilişki varsa: w11 > w12 > w22, nesiller sonra canlı popülasyonunun nasıl değişmesini beklersiniz?
Sorudan görülebileceği gibi en yüksek uyum başarısı homozigot baskın olana aittir. Baskın genotipin her zaman en yüksek uyum başarısına sahip olması gerekmediğini önemle hatırlayınız. Genetik baskınlık ile, popülasyon içi uyum başarısı arasında hiçbir ilişki yoktur. Soruya dönecek olursak: sayısız nesil geçtikten sonra, kişi ilk etapta homozigot çekinik (22) olanların hepsinin elenmesini beklediğini söyleyebilecektir. Çünkü en düşük uyum başarısı ona aittir ve kademeli olarak yok olacaktır. Ancak aynı zamanda heterozigot bireylerin (12) de uyum başarısı homozigot baskından (11) düşüktür. Dolayısıyla eğer ki çevre şartları değişmezse ve her zaman bu ilişki korunursa, 3 genotipten sadece 1 tanesi hayatta kalacaktır: homozigot baskın olan (11). Diğer ikisi de elenip yok olacaktır, çünkü uyum başarıları daha düşüktür.
Elbette gerçek hayatta çevre sürekli değişir. Uyum başarısı dinamik bir yapıdır ve her an değişmektedir. 1 yıl önce uyumsuz olan bir genotip, 1 yıl sonra en uyumlu hale gelebilir. Benzer şekilde, gerçek hayatta sürekli mutasyonlar, crossing-over, vb. çeşitlilik mekanizmaları yeni genetik kombinasyonlar yaratmaktadır. Burada sadece tek bir adet gen varmış gibi düşündüğümüzü hatırlayınız. Ancak normalde bir canlıyı tanımlayan on binlerce gen vardır ve her birinin bireye kattığı uyum başarısı birbirinden farklı olacaktır. Bunların dengesi sayesinde türler bu kadar tekdüze bir şekilde evrimleşip yok olmazlar. Ancak eğer ki tüm genler göz önüne alındığı zaman, 3 farklı özellikteki popülasyon arasında böyle bariz bir ilişki varsa, sadece 1 tanesinin uzun vadede hayatta kalacağını görebiliriz.
İşte bu da bizi çok ilginç bir gerçeğe götürür: gerçekten de evrimsel süreçte genellikle birbirine yakın canlı türlerinden sadece 1 ya da birkaç tanesi hayatta kalabilir. Örneğin Homo cinsine ait 14 civarında farklı tür olmasına ve bunların 4-5 tanesi son birkaç yüz bin yılda hayatta olmasına rağmen, günümüze sadece bizler, yani Homo sapiens erişebilmiştir; diğer tüm insan türleri yok olmuştur. Bu sadece insanda görülen bir şey değildir; sayısız türde böyledir. Çünkü farklı türlerin uyum başarıları arasında farklar vardır ve bu farklar, er ya da geç bir çoğunun yok olmasına ve bazılarının hayatta kalmasına neden olur. Zaten evrim de budur. Ancak bu süreçte, yeni türleşmeler ve değişmeler meydana gelerek, diğerlerine göre başarılı olan popülasyonlar içerisinde de birkaç tane daha da başarılı soy hattı oluşabilir. İşte Evrim Ağacı'nın dalları da bu şekilde çatallanarak büyümeyi sürdürür, yeni türler böyle var olur.
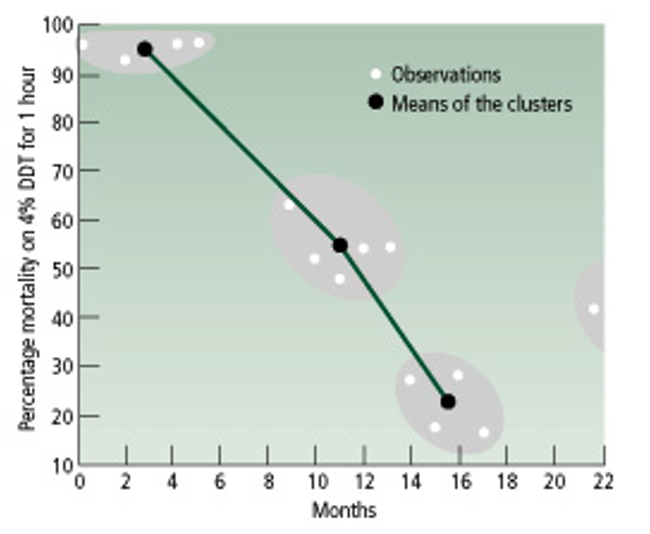
Yukarıdaki grafik, bunun deneysel örneklerinden birine aittir. Anofel tipi sivrisinekler olarak bilinen Anopheles culicifacies türü sivrisineklerle mücadele etmek için %4 oranında DDT kimyasalı 1 saat boyunca sıkılmıştır. Sıkımın başladığı 0 noktasından sonraki ilk 4-5 ayda ölüm oranları, grafikte gösterildiği gibi %98 dolaylarında seyretmiştir. Yani ilaç, her 100 sivrisinekten 98'ini öldürebilecek kadar güçlüdür. Ancak her sivrisinek birbiriyle aynı özellikte değildir. Çeşitlilik mekanizmalarından ötürü bazıları diğerlerinden daha dirençlidir ve bu kimyasala direnerek hayatta kalabilirler. İşte bunlar, ilk etapta sadece %2'lik, ufacık bir kısımdır.
Ancak bunların üremeye başlamasıyla, hızla dirençli popülasyonlar evrimleşmiştir. Görülebileceği gibi, bu sebeple aylar içerisinde ölüm oranları hızla 0'a doğru düşmeye (çakılmaya) başlamıştır. Sadece 10-11 ay sonunda, birebir aynı ilaç, aynı bölgeye sıkıldığında, aynı sivrisineklerden evrimleşmiş yeni popülasyonun sadece %55 civarını öldürmeyi başarabilmiştir. Başlangıçtan 16 ay sonra ise ölüm oranları %23'e kadar düşmüştür. Açık bir şekilde sivrisinekler, ortamdaki öldürücü kimyasala direnç kazanacak şekilde evrim geçirmektedirler. Bu direnç kazanımı, hem genlerinin, hem de uzun vadede fiziksel görünümlerini tetikleyecek değişimlerle mümkün olmaktadır. Bu tür olan, belli bir yöne doğru tüm popülasyonun evrimleşmesine neden olan seçilim türüne Yönlü Doğal Seçilim dendiğini hatırlayınız.
Ancak unutmayınız ki eğer ki ilk popülasyonda yeterli çeşitlilik olmasaydı ve hepsi ölseydi, böyle bir evrim asla mümkün olamayacaktı. Yani sivrisinekler "ihtiyaçtan ötürü" veya "bilerek" evrim geçirmediler. Hayatta kalanların üremesi, ister istemez evrime neden oldu. Zaten evrimsel değişimler her zaman bu şekilde olur. Ne var ki neredeyse her zaman popülasyonlarda sayısız çeşitli ortama farklı tepkiler verebilecek kadar geniş bir çeşitlilik söz konusudur. Evrim, genellikle üzerine etkiyecek çeşitlilik bulmakta zorlanmaz (çeşitlilik mekanizmaları sağ olsun).
Ekleyici Yönlü Seçilim
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F57673223_additivepng.png)
Yukarıdaki grafikte 3 farklı fare genotipi görülmektedir. Görebileceğiniz gibi BB genotipine sahip olanlar tamamen siyah, TT genotipine sahip olanlar ise tamamen turuncudur. BT genotipine sahip olup, heterozigot olanlarsa siyah-turuncu bir renge sahiptirler. Bu, eşbaskınlık durumuna bir örnek olarak görülebilir. Yani iki alel birbirine eşit derecede baskındır. Böyle bir farenin fotoğrafını yazının başında göstermiştik.
Belli bir ortamda, BB genotipinin göreceli uyum başarısı 1 olarak verilmiştir. BT genotipi 0.8, TT genotipi ise 0.6 değerine sahiptir (sol taraftaki grafik). Hatırlayacak olursanız, seçilim baskısı göreceli uyum başarısının 1'den çıkarılmasıyla bulunuyordu. Çünkü bir canlı ne kadar uyumluysa, üzerindeki seçilim baskısı o kadar küçük olacaktır (ve tam tersi). Dolayısıyla 3 genotipin de üzerine etki eden seçilim baskısını (s ile ifade ediyoruz) sırasıyla 0 (hiç yok), 0.2 ve 0.4 olarak hesaplayabiliriz. Ayrıca bu durumdaki ortalama uyum başarısını da şu şekilde hesaplayabiliriz:
ῶ = (0.25 x 1.0) + (0.5 x 0.8) + (0.25 x 0.6)
ῶ = 0.8
Görülebileceği gibi bu durumda ortalama uyum başarısı, tam olarak heterozigotunkine (BB'nin başlangıçtaki uyum başarısına) eşittir. Ancak bu, heterozigotun uyum başarısının iki homozigotun uyum başarılarının tamı tamına ortasında olmasındandır. Normalde her zaman böyle olmayabilir; bu durumları az sonra göreceğiz.
Uyum başarıları veya seçilim baskıları bu şekilde dağılan genetik özelliklere bağlı meydana gelen yönlü seçilime Ekleyici Yönlü Seçilim adını veriyoruz. Çünkü heterozigot olan bireyler, iki uçtaki homozigot bireylerin ortasında bir uyum başarısına (ve seçilim baskısına) sahiptir. Yani adeta iki genotipin uyum başarıları (ve seçilim baskıları) birbirine eklenip, ortalamaları alınarak heterozigot genotipi yaratmaktadırlar.
Bu durumda, sağ taraftaki grafiğe ulaşırız: en yüksek uyum başarısı (en düşük seçilim baskısı) BB genotipinde olduğu için, alel frekansı nesiller geçtikçe ondan tarafa kayacaktır. Yani popülasyon içerisinde sürekli olarak BB bireylerin sayısı artacak, BT ve TT bireylerin sayısı azalacaktır. Bu süreç, ya popülasyondaki bireylerin frekansı 1'de (veya 0'da) sabitlenene kadar, ya çevre ve seçilim baskısı değişene, ya da artık etki edecek bir çeşitlilik kalmayıncaya kadar devam eder. Ancak tekrar söylemek gerekirse, genelde bu sabitlenmeler gerçekleşmeden önce popülasyon veya çevre değişir, böylece evrimin gidişatı da yön değiştirir.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F58308242_frekapng.png)
Ekleyici Yönlü Seçilim dolayısıyla, yukarıda anlattığımız gibi B alelinin frekansı sürekli artar. Eğer ki çevre değişmezse, sonunda %100'e (1'e) ulaşarak sabitlenir. Ancak bu sırada uyum başarısı da sürekli artar. Çünkü ortalama uyum başarısının tanımı gereği alellerin frekansı ile onların uyum başarılarının çarpımlarının toplamı olduğunu hatırlayın. Eğer ki B'nin uyum başarısı en yüksekse ve süreç içerisinde bu sebeple frekansı sürekli olarak artıyorsa, o popülasyonun uyum başarısı da sürekli olarak artar.
Baskın Yönlü Seçilim
Yukarıda da izah ettiğimiz gibi, heterozigotların uyum başarısı her zaman homozigotların tam ortasında olmayabilir. Örneğin kimi durumda heterozigotlar da, homozigot baskınlar kadar yüksek uyum başarısına sahip olabilirler. Bu durumda eş baskınlık değil, tam baskınlık durumu görülür. Dolayısıyla BT aleline sahip olan heterozigotlar, iki alelin bir karışımını değil, baskın alelin özelliklerini gösterir. Dolayısıyla BB genotipinin de, BT genotipinin de uyum başarısı eşit olacaktır. Çünkü genellikle evrim, genotipin yarattığı fenotip üzerine etki eder. Bu durumda grafikler aşağıdaki gibi değişir.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F59612404_domipng.png)
Görülebileceği gibi tek değişen heterozigotun uyum başarısının 0.8 yerine, tıpkı BB genotipi gibi 1 olmasıdır. Bu durumda ortalama uyum başarısı da 0.8'den yukarı kayacaktır:
ῶ = (0.25 x 1) + (0.5 x 1) + (0.25 x 0.6)
ῶ = 0.9
Tam da beklediğimiz gibi... Ayrıca sağ taraftaki grafiğin de yukarı doğru eğrildiğine dikkat ediniz. Böylece popülasyondaki seçilim süreci çok daha çabuk tamamlanabilir ve sabitlenir.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F60328417_grupng.png)
Buradaki kilit nokta, uyum başarısını düşüren T alelinin ne olursa olsun seçilimden kurtulmayı başarmasıdır. İşte bu sebeple çekinik alelleri evrimin tamamen silmesi çok zordur. Heterozigotlar içerisinde korunarak çekinik aleller popülasyonda kalmayı başarırlar. Bir diğer deyişle çokbiçimlilik (polimorfizm) korunmuş olur. Bir genin farklı alelleri silinemeden kalır.
Çekinik Yönlü Seçilim
Yukarıdaki durumun tam tersi de söz konusu olabilir. Bu durumda heterozigot olan bireyler, çekinik homozigotlara daha yakın uyum başarısına sahip olabilirler. Eğer ki çekiniklerin uyum başarısı daha düşükse, ortalama uyum başarısı da 0.8 olarak verdiğimiz (durumdan duruma değişebilir, bu bir örnektir) ortalama değerden daha düşük olmalıdır. Bu durumu görelim:
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F60703837_dominpng.png)
Bu durumda ortalama uyum başarısını hesaplarsak:
ῶ = (0.25 x 1) + (0.5 x 0.6) + (0.25 x 0.6)
ῶ = 0.7
Yine tam beklediğimiz gibi...
Bu durumda evrimin uyum başarısını arttırma hızı da düşer. Bu sebeple başlangıçtaki değişim öncekilere göre daha yavaş olur. Ancak yine de nihayetinde B aleli 1'e sabitlenecektir; çünkü halen en yüksek uyum başarısı ona aittir. Bu grafiği görecek olursak:
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fdosyalar%2Ficerikler%2F60882193_bbpng.png)
İşte farklı genotiplerin farklı uyum başarıları evrimi bu şekilde farklı yönlere sürükleyebilir. Bahsettiğimiz gibi, burada bariz bir uyum başarısı farkı söz konusudur ve evrim her zaman uyum başarısını maksimuma çıkarmaya çalışır; daha doğrusu doğanın yasası budur ve bu yasayı biz evrim olarak isimlendiririz. Ancak daha sonraki yazılarımızda göreceğimiz gibi, her zaman doğada böyle doğrusal bir uyum başarısı dağılımı bulunmaz. Dolayısıyla bazı başka faktörler de evrimin hızını ve yönünü etkileyebilir, tamamen değiştirebilir.
Ancak bunları sonraki yazılarımıza saklayacağız.
Evrim Ağacı'nda tek bir hedefimiz var: Bilimsel gerçekleri en doğru, tarafsız ve kolay anlaşılır şekilde Türkiye'ye ulaştırmak. Ancak tahmin edebileceğiniz gibi Türkiye'de bilim anlatmak hiç kolay bir iş değil; hele ki bir yandan ekonomik bir hayatta kalma mücadelesi verirken...
O nedenle sizin desteklerinize ihtiyacımız var. Eğer yazılarımızı okuyanların %1'i bize bütçesinin elverdiği kadar destek olmayı seçseydi, bir daha tek bir reklam göstermeden Evrim Ağacı'nın bütün bilim iletişimi faaliyetlerini sürdürebilirdik. Bir düşünün: sadece %1'i...
O %1'i inşa etmemize yardım eder misiniz? Evrim Ağacı Premium üyesi olarak, ekibimizin size ve Türkiye'ye bilimi daha etkili ve profesyonel bir şekilde ulaştırmamızı mümkün kılmış olacaksınız. Ayrıca size olan minnetimizin bir ifadesi olarak, çok sayıda ayrıcalığa erişim sağlayacaksınız.
Makalelerimizin bilimsel gerçekleri doğru bir şekilde yansıtması için en üst düzey çabayı gösteriyoruz. Gözünüze doğru gelmeyen bir şey varsa, mümkünse güvenilir kaynaklarınızla birlikte bize ulaşın!
Bu makalemizle ilgili merak ettiğin bir şey mi var? Buraya tıklayarak sorabilirsin.
Soru & Cevap Platformuna Git-
 7
7
-
 7
7
-
 6
6
-
 5
5
-
 2
2
-
 1
1
-
 1
1
-
 0
0
-
 0
0
-
 0
0
-
 0
0
-
 0
0
- J. Herron. (2019). Evolutionary Analysis. ISBN: 0321616677. Yayınevi: Pearson.
- D. Futuyma. (2013). Evolution, Douglas Futuyma. ISBN: 1605351156. Yayınevi: Oxford University Press.
Evrim Ağacı'na her ay sadece 1 kahve ısmarlayarak destek olmak ister misiniz?
Şu iki siteden birini kullanarak şimdi destek olabilirsiniz:
kreosus.com/evrimagaci | patreon.com/evrimagaci
Çıktı Bilgisi: Bu sayfa, Evrim Ağacı yazdırma aracı kullanılarak 09/02/2026 13:08:05 tarihinde oluşturulmuştur. Evrim Ağacı'ndaki içeriklerin tamamı, birden fazla editör tarafından, durmaksızın elden geçirilmekte, güncellenmekte ve geliştirilmektedir. Dolayısıyla bu çıktının alındığı tarihten sonra yapılan güncellemeleri görmek ve bu içeriğin en güncel halini okumak için lütfen şu adrese gidiniz: https://evrimagaci.org/s/398
İçerik Kullanım İzinleri: Evrim Ağacı'ndaki yazılı içerikler orijinallerine hiçbir şekilde dokunulmadığı müddetçe izin alınmaksızın paylaşılabilir, kopyalanabilir, yapıştırılabilir, çoğaltılabilir, basılabilir, dağıtılabilir, yayılabilir, alıntılanabilir. Ancak bu içeriklerin hiçbiri izin alınmaksızın değiştirilemez ve değiştirilmiş halleri Evrim Ağacı'na aitmiş gibi sunulamaz. Benzer şekilde, içeriklerin hiçbiri, söz konusu içeriğin açıkça belirtilmiş yazarlarından ve Evrim Ağacı'ndan başkasına aitmiş gibi sunulamaz. Bu sayfa izin alınmaksızın düzenlenemez, Evrim Ağacı logosu, yazar/editör bilgileri ve içeriğin diğer kısımları izin alınmaksızın değiştirilemez veya kaldırılamaz.
:sharpen(0.5,0.5,true)/evrimagaci.org%2Fpublic%2Fimages%2Flogo-50.png)